HYPOTHALAMUS and NEUROENDOCRINE SYSTEMS
FOUNDATIONS (
2. Major Fiber Systems of the Hypothalamus
4. Hypothalamic Nuclei
5. Magno- and Parvocellular Neurosecretory System
6. Reflex Control of Vasopressin and Oxytocin Secretion
11. Behavioral State Control
12. Drinking
13. Temperature Regulation
14. Further reading
15. Legend to figures
The
hypothalamus control autonomic, behavioral and neuroendocrine functions as
summarized in Figure 1, 2, 3.
1. Boundaries and Subdivisions (Figures 4-17)
The hypothalamus forms the ventral part of the
diencephalon. The hypothalamus can be divided
longitudinally into periventricular, medial and lateral cell groups. The medial and periventricular
hypothalamus contains most of the neurons concerned with regulation of the
pituitary, but also important efferent sources for projections to brainstem and
spinal autonomic areas. The medial
hypothalamus has, in addition, extensive reciprocal connections with the
medial division of the 'extended amygdala’.
The hippocampus, either directly or via the septum, also sends afferents
to medial hypothalamus. The lateral
preoptic-hypothalamic (LPO-LH) continuum
contains numerous cells, which are interspersed among fibers of the medial
forebrain bundle (MFB). The LPO-LH area shares a wide variety of reciprocal
connections with the forebrain, caudal brainstem, and spinal cord. The physiology of this area is complicated by
the fact that many axons traverse this area, which may or may not synapse
locally.
2. Major Fiber Systems of the Hypothalamus (Figures 18-25)
Some of the heavily myelinated hypothalamic fiber
tracts, e.g. fornix, mamillothalamic tract, stria medullaris, stria terminalis,
medial forebrain bundle can be identified by blunt dissections or using myelin
staining, however, the direction of fibers within these tracts can be
identified only by experimental tract-tracing methods.
Fornix. The fornix connects the hippocampal formation with the
septal area, anterior thalamus and hypothalamus (Figure 19).
Mammillothalamic Tract and Mammillary Peduncle.
A capsule of heavily myelinated fibers surrounds the mammillary body in
the caudal part of the hypothalamus. Its
function is not well known. Most of its
efferent fibers leave the mammillary body in a dorsal direction as the
mammillothalamic tract, which proceeds towards the anterior thalamic
nuclei. Collaterals of the
mammillothalamic fibers form the mammillotegmental tract, which projects to
tegmental cell groups in mesencephalon.
These cell groups in turn give rise to the mammillary peduncle, which
terminates primarily in the lateral mammillary nucleus (Figure 21).
Stria Medullaris. The stria
medullaris, which can be easily recognized on the mediodorsal side of the thalamus,
connects the lateral preoptic-hypothalamic region with the habenular
complex. However, like most other
hypothalamic pathways, the stria medullaris is a complicated bundle that
contains many different fiber components with various origins and terminations
(Figure 21).
Stria Terminalis. The stria
terminalis reciprocally connects the amygdaloid body and the medial
hypothalamus. Similar to the fornix, the
stria terminalis makes a dorsally convex detour behind and above the
thalamus. It can be identified in the
floor of the lateral ventricle, where it accompanies the thalamostriate vein in
the groove that separates the thalamus from the caudate nucleus. In the region of the anterior commissure, the
stria terminalis divides into different components, which distribute their
fibers to the bed nucleus of the stria terminalis, medial hypothalamus and
other areas in the basal parts of the forebrain. The stria terminalis is an important pathway
for amygdaloid modulation of hypothalamic functions. The amygdaloid body is also related to the
lateral hypothalamus through a diffuse ventral amygdalofugal pathway
that spreads out underneath the lentiform nucleus (Figure 22).
Dorsal Longitudinal Fasciculus The DLF is a component of an
extensive periventricular system of descending and ascending fibers that
connects the hypothalamus with the midbrain gray and other regions in the pons
and medulla oblongata including preganglionic autonomic nuclei.
Medial Forebrain Bundle. The MFB
is an assemblage of loosely arranged; mostly thin fibers, which extends from
the septal area to the tegmentum of the midbrain. It traverses the lateral preoptico-hypothalamic
(LPO-LH) area, the scattered neurons of which are collectively designated as
the bed nucleus of the MFB. The bundle
is highly complex, comprising a variety of short and long ascending and
descending links (Figure 18).
3. Connections of the Hypothalamus
Most of the connections of the
hypothalamus consist of fine, unmyelinated fiber systems that cannot be traced
accurately in normal myelin- or fiber-stained preparations. As a result, much of what is now known about the
connections of the hypothalamus has been learned in the last decade or so,
since the introduction of the axonal tracer methods. These connections are summarized below.
Afferents
Cortical Inputs. Cortical inputs
to the hypothalamus in the rat arise primarily from insular, lateral frontal,
infralimbic, and prelimbic areas. These
afferents principally supply the lateral hypothalamic area.
Visceral inputs. Viscerosensory
information reaches the hypothalamus via ascending projections of the nucleus
of the solitary tract (NTS), that receives input from the major visceral organ
by way of the glossopharyngeal (IX) and vagal (X) cranial nerves. The NTS is
the first region in the CNS that process information about visceral,
cardiovascular, respiratory functions as well as taste. In the monkey and
human, presumably the visceral afferent influence from the NTS is relayed to
the hypothalamus via the projection of the NTS to the parabrachial
nucleus. Neurons in the paraventricular
hypothalamic nucleus and the lateral hypothalamic area receive direct
(synaptic) input from the NTS.
Olfactory inputs. In rodents,
olfactory input arrives via relays in the olfactory tubercle, anterior
olfactory nucleus, corticomedial amygdala and olfactory cortex. From these regions, secondary olfactory
afferents terminate throughout the lateral hypothalamus.
Visual inputs may reach the hypothalamus via a direct retinal
projection. In all mammalian species,
including humans, some retinal fibers leave the optic chiasm and pass dorsally
into the hypothalamus, where they innervate the suprachiasmatic nuclei, the
endogeneous circadian clock.
Somatosensory information may also reach the hypothalamus via a direct
route: a projection to the lateral hypothalamic area from wide-dynamic-range
mechanoreceptive neurons in the spinal dorsal horn.
Auditory input. Despite extensive study, no
direct projection to the hypothalamus from the auditory system has been
identified. Recently, however, it has
been shown that acoustic stimulation induce LH release in birds (MeiFang et
al., 1998). Many hypothalamic neurons respond best to complex sensory stimuli,
suggesting that the sensory information that drives them is highly
processed. It is likely, therefore, that
much of the sensory information that reaches the hypothalamus travels by
polysynaptic routes involving convergence of cortical sensory pathways in the
amygdala, hippocampus and cerebral cortex.
Monoamine cell groups. Each of
the classes of monoamine cell groups in the rat brainstem provides innervation
to the hypothalamus.
Projections from limbic regions. Hippocampal efferents via the
precommissural fornix-lateral septum innervate all three longitudinally
organized columns of the hypothalamus. A distinct subdivision of the
hippocampus, the subiculum, project through the postcommissural fonix to the
mammillary bodies. Several cell groups of the amygdala project via the stria
terminalis or the ventral amygdalofugal pathway to the hypothalamus. The
ventral subiculum project via the medial corticohypothalamic tract to the
medial hypothalamic cell groups.
The Circumventricular Organs (CVOs). Chemosensory information
from plasma (blood-borne molecules) or CSF reaches the hypothalamus via input
from projections of CVOs. CVOs has specialized fenestrated capillaries,
permitting relatively large molecules to leave the vascular bed and enter the
extracellular milieu. Two of these regions, the subfornical organ (SFO) and
area postrema (AP) have extensive connections with hypothalamic nuclei involved
in neuroendocrine and homeostatic regulation. Two other CVOs, the organon
vasculosum laminae terminalis (OVLT) and the median eminence (ME), are located
within the hypothalamus.
Efferents
The main outflow of hypothalamic nuclei are
directed 1) median eminence
(parvocellular neurons) (Figure 24), 2) posterior pituitary
(magnocellular) (Figure 26)
to influence neuroendocrine responses; 3) sympathetic and parasympathetic pregangionic cell groups
in the brainstem and spinal cord (Figure
31) to influence autonomic functions (primarily originating in
the dorsal, medial and lateral parvocellular division of the PVN); 4) several cell groups in the
hypothalamus project to the amygdala, bed nucleus of the stria
terminalis, to the basal nucleus of Meynert, periaqueductal gray
(PAG), visceral sensory areas of the thalamus (ventroposterior
parvocellular nucleus) cerebral cortex (anterior insular cortex,
anterior tip of the cingulate cortex), and brainstem (NTS, parabrachial nucleus) (Figure 25) to influence various behavioral
responses.
4. Hypothalamic Regions (nuclei, areas) (Figs.
6, 12, 13, 14, 15)
Four lines of evidence support
the view that the suprachiasmatic nucleus (SCN) is the dominant
mammalian endogeneous timekeeper. 1) This
nucleus receive afferents directly (retinohypothalamic tract) and indirectly
(via the LGN) from the retina in order to synchronize otherwise free-running
circadian rhythms with the day-night cycle.
2) Lesions of the SCN typically alter only the temporal organization of
a function (see later), the function itself is not changed. 3) Isolation of the
SCN either in vitro or in vivo, does not alter its ability to generate
circadian signal. 4) Transplantation of a fetal SCN into the third ventricle of
arrhythmic hosts with SCN lesions restores circadian rhythm with a period that
reflects donor, not host, rhythm (
The paraventricular nucleus (PVN), in addition to
the magnocellular vasopressin and oxytocin neurons, contains several subgroups
of small (parvicellular) neurons
containing a variety of
putative neurotransmitters. Some of the
parvicellular neurons (e.g. CRF=corticotropin releasing factor) project to the
median eminence where they participate in the regulation of the anterior
pituitary. Other neurons in
the PVN project to sympathetic and
parasympathetic autonomic areas
in the medulla
and the intermediolateral cell
columns of the spinal cord. The PVN has
been implicated in a variety of behaviors including feeding, thirst,
cardiovascular mechanisms as well as organization of autonomic and endocrine
responses to stress.
The subparaventricular zone (SPVZ) is thought to
play a role in amplifying circadian output from the SCN
The supraoptic nucleus (SON) contains vasopressin
and oxytocin and project with similar axons originating in the PVN to the
posterior pituitary.
The anteroventral third ventricle region (AV3V) is a term that encompasses several preoptic subnuclei and the OVLT that is important in osmo-and volum regulation.
The ventrolateral preoptic area (VLPO) is a
recently coined term to define cells that are sleep-active.
The arcuate nucleus (ARC)
among others contains dopamine, which acts as a prolactin-inhibiting factor at
the median eminence. In additions, its
neurons are estrogen sensitive and project to the preoptic LHRH neurons. This circuit is involved in the regulation of
gonadotropin secretion and sexual behavior during female reproductive cycle.
The ventromedial nucleus
(VMH) in addition to its output to the median eminence, with their other
projections is thought to participate in the organization of reproductive
behavior, as well as in metabolic regulatory functions.
The dorsomedial hypothalamic nucleus (DMH) among others is involved in mediating leptin actions to the PVN. Fibers from the SCN via the DMH towards the locus coerules are suggested to participate in circadian regulation of sleep and waking.
The tuberomammillary nucleus (TMN) is located in the caudoventral part of the lateral hypothalamus. Its neurons contain the sleep-active histamin projection system.
The mammillary body is at
the caudal border of the hypothalamus.
The lateral and medial mammillary nuclei are the recipient of a massive
input from the hippocampus that arrives via the fornix. These nuclei project via the mammillo-thalamic
tract to the anterior nuclei of the thalamus.
These nuclei are frequently damaged in Korsakoff's patients.
The lateral hypothalamic area
(LHA) contain neurons scattered around the medial forebrain bundle
The perifornical area (PFA)
consists of cells around the hypothalamic course of the fornix. Two important
cell populations are discussed in this chapter: 1) orexin/hypocretin cells and
2) melanin-concentrating hormone cells (MCH)
5. Magno- and Parvocellular Neurosecretory System (Figures 26-33)
The
magnocellular neurons of the supraoptic
(SON) and paraventricular (PVN)
nuclei along with scattered clusters of large cells between these two nuclei
comprise the hypothalamo-hypophyseal system. These cells send oxytocin and vasopressin
containing fibers to the posterior pituitary where these substances are
released into the peripheral circulation.
Vasopressin is the well-known antidiuretic hormone (ADH) and is
released in response to changes in the osmotic pressure of circulating blood or
extracellular space. ADH controls the
water-balance. In particular, it is
responsible for the retention of water, which is regulated by the effect of
vasopressin on the distal tubules of the kidneys.
Oxytocin,
through its effect on the uterine smooth muscle and the myoepithelial cells of
the mammary glands, promotes uterine contraction during birth and milk ejection
after birth. Potent stimulatory input
for uterine contraction reaches the brain via afferents from the vagina or
cervix and the nipples.
Hypothalamic (parvocellular) neurons originating in the preoptic, arcuate, ventromedial, periventricular, paraventricular nuclei transport a variety of releasing and inhibiting hormones to the portal vessels of the median eminence. Fenestrated capillaries loop through the median eminence coalesce to form long portal vessels that travel along the infundibular stalk where they are continuous with vascular sinuses in the anterior pituitary. These substances are then transported to the capillary beds of the anterior pituitary where they regulate the secretion of the pituitary troph hormones: TRH (Thyrotropin-Releasing Hormone) → TSH (Thyrotropin), CRH or CRF (Corticotropin-Relasing Hormone) → ACTH (Adrenocorticotropin Hormone), GnRH (Gonadotropin-Releasing Hormone) → FSH (Follicle-Stimulating Hormone) and LH (Luteinizing Hormone), GHRH (Growth Hormone-Releasing Hormone) and GHRIH (somatostatin) → GH (Growth Hormone), PRF (Prolactin-Releasing Factor) and PIF (Prolactin Release-Inhibiting Factor=dopamine) → Prolactin, MRF (Melanocyte-stimulating hormone Releasing Factor) and MIF (Melanocyte-stimulating hormone release Inhibiting Factor) → MSH (Melanocyte-Stimulating Hormone). Figure 3 summarizes the target organs upon which the pituitary troph hormones act. Figures 26 summarizes the design of the parvo- and magnocellular neurosecretrory system. Figures 27-28 details aspect of organization of the median eminence-arcuate nucleus region and Figure 29 shows the relationship of troph-hormone producing cells to fenestrated capillaries in the anterior pituitary.
The magno- and parvocellular cell groups producing
the hypothalamic hormones receive a variety of stimuli from different parts of
the brain, primarily within the hypothalamus, but also from extrahypothalamic
areas including the amygdaloid body, hippocampus and various brainstem areas (Figures
31, 33). Furthermore, it is well known that monoamines
and several neuropeptides serve as modulators of the neuroendocrine system, and
both monoaminergic and peptidergic fibers, besides those carrying the specific
hypothalamic hormones, can be traced to the periventricular zone and even into
the median eminence, where they would have an opportunity to interact with the
parvicellular neurosecretory system or even discharge neuroactive substances
directly into the portal system.
The
subject of neuroendocrine control mechanism is complicated further by the fact
that many neurons in the nervous system, including the hypothalamic
magnocellular and parvocellular neurosecretory neurons contain two or even
several neuroactive substances. A
well-known example is provided by the parvocellular CRF neurons in the
PVN. They also contain vasopressin and
the two substances are released together into the portal vessels, through which
they are likely to cooperate in the control of ACTH-release from the adenohypophysis.
Hypothalamic neurons, including the neurosecretory neurons, are also subject to
hormonal feedback control. Such feedback
mechanisms are often quite complicated in the sense that they involve not only
the neurosecretory hypothalamic neurons but also hormone sensitive cells in
other brain regions, which in turn are in a position to modulate
hypothalamo-hypophysial function. Peripheral hormones (e.g. estrogen, etc)
exert their feedback actions also at the level of the median eminence and the
anterior pituitary.
6. Reflex Control of Vasopressin and Oxytocin Secretion (Figures 34, 35, 36, 37, 38)
The nonapeptides, oxytocin (OT) and vasopressin
(VP), two major biologically active hormones, are synthesized in separate cell
populations in the supraoptic and paraventricular nuclei of the
hypothalamus. These peptides are carried
by axoplasmic transport to various areas within the CNS and to the posterior
pituitary. OT and VP are released from
nerve endings in the neural lobe of the pituitary to reach the systemic
circulation and influence primarily fluid balance (VP) and milk ejection/uterus
contraction (OT). In addition, by their
axonal projections in the CNS, VP and OT also play a role in neurotransmission.
Figure 32 shows the
distribution of vasopressin and oxytocin neurons in the PVN and SON.
The vasopressin gene encode a 145 amino acid
prohormone that is packaged into neurosecretory granules of the magnocellular
neurons. During axonal transport of the granules from the hypothalamus to the
posterior pituitary, enzymatic cleavage of the prohormone generates the final
products: VP, neurophsyin and a carboxy-terminal glycoprotein. When afferent
stimulation depolarizes the VP-containing neurons, the three products are released
into capillaries of the posterior pituitary. Peripheral VP functions largely to
maintain arteriolar perfusion pressure and intravascular volume. One of the most potent effective stimuli
for VP secretion is a rise in extracellular osmolality. Although less potent, other indicators of
extracellular fluid depletion also stimulate VP release, including decreased
plasma volume (hypovolemia, hemorrhage), decreased blood pressure
(hypotension), and peripheral hypoxia or hyperkapnia or both. In contrast,
drinking fluids, even when they are hypertonic, results in an abrupt fall in
plasma VP levels, presumably via stimulation of osmoreceptors in the
oropharynx. In addition, various
stressors, fever, pain and nausea and emetic agents such apomorphine causes VP
(and OT) release (Figure 34).
Effectors.
Circulating VP maintains extracellular fluid balance by acting a) at the kidney,
where it stimulates (through VP receptors) increased retention of water and
enhanced Na and Cl excretion, b) at arterioles, where it is one of the
most potent vasoconstrictors, c) it also modulate sympathetic transmission
and d) affect the baroreceptor reflex by a central effect mediated by
the area postrema. Peripheral VP has also been found to have effects on hepatic
glycogenolysis, platelet aggregation and blood coagulation. Under emergency
conditions, the peptide causes vasoconstriction in skin, gastrointestinal tract
and kidney. It serves to shunt blood
from these tissues to the brain, heart and lung (Figure 37).
Peripheral Osmo- Chemo Receptors and Pathways. Vasopressin
is under tonic inhibitory influence both from atrial receptors of the
heart (low pressure receptors) and from baroceptors (high pressure) in
the aortic arch (X) and carotid sinus (IX).
Reduction in the discharge of these receptors by a decrease in blood
volume or blood pressure results in the release of vasopressin. An additional excitatory influence on vasopressin
release is provided by carotid body chemoreceptors and peripheral osmo-
and stretch[pressure]-receptors. The
signal from arterial baroreceptors, cardiopulmonary receptors and peripheral
osmoreceptors (in the mesenteric and hepatic vasculature) is carried through
the IX and X nerves to the NTS. From the
NTS information though GABAergic neurons or directly may reach Al noradrenergic
neurons which project to SON and PVN VP neurons to stimulate VP release. Bilateral carotid occlusion releases vasopressin
and leads to activation of carotid body chemoreceptors. It is not clear, however, how the excitatory
input from the chemoreceptors reaches the SON or PVN. A possibility is that excitatory input to the
VP neurons may arise from cholinergic neurons that lie dorsal to SON. Indeed, ACh injected into the SON results in
nicotine mediated vasopressin release (Figures 35, 36).
Humoral Factors (Angiotensin II) and Pathways. Lesion
studies established that both the subfornical organ (SFO) and the AV3V
(anteroventral 3rd ventricle region which includes various preoptic
nuclei and the OVLT) are involved in the neural regulation of salt and water
balance. Both the SFO and the AV3V
regions is the major target for the dipsogenic action of blood born Ang
II. When blood pressure falls, the
kidneys release renin into the bloodstream. Renin triggers a biochemical
cascade that produces AII (angiotensin) (Figure 38). Ang II is released from the periphery as a
chemical signal of hypovolemia and hypotension.
Ang II stimulates VP secretion by acting at the SFO. Because SFO lacks
blood-brain barrier, the AII receptors can detect very small increases in blood
level of AII. Additional evidence suggests that the SFO and OVLT and the
magnocellular neurons themselves may be sensitive to changes in extracellular
osmolality and sodium concentration.
Afferents from the SFO and AV3V reach both PVN and SON neurons. The
projection from SFO to PVN and SON use ANG II as transmitters.
Additional osmo- or Na receptors are located in
the area postrema that project to the NTS.
NTS afferents reach the AV3V region also through the parabrachial
area. Afferents from the bed nucleus of
the stria terminalis to PVN, SON serves to integrate central cardiovascular and
"limbic" information.
Clinicopathology. The
lack of VP results in a condition known as diabetes insipidus (DI,
In contrast, circulating OT is best known in
female reproduction, where it is involved in the maintenance of parturition and
the initiation of lactation. Thus, OT
acts on the smooth muscle of the endometrium during labor and delivery to
increase the frequency and intensity of uterine contractions, and on the
myoepithelial cells surrounding mammillary alveolar glands to cause milk
let-down in response to suckling. An
increase in circulating OT also accompanies ejaculation in males. OT also stimulates the release of both
insulin and glucagon from the pancreas, and act on adipocytes, indicating that
it plays a role also in metabolic regulation related to feeding. Vaginal and uterine distension receptors,
somatic sensory receptors from the nipple and breast and nociceptive
information from much of the body are all relayed initially to the dorsal horn
of the spinal cord, from where axons project to the Al cell group and the
caudal NTS. From here, ENK, SS and
inhibin B pathways may mediate specific stimuli to OX neurons in the
hypothalamus.
7. Brain-Pituitary Gonadal Axis (Figures 39, 40, 41)
The hypothalamus
plays a major integrative role in the control of maternal and reproductive
behavior, including sexual development, and differentiation, as well as sexual
behavior. Important stimuli for the
various aspects of reproductive functions come from a variety of exteroceptive
and interoceptive sources including circulating gonadal steroids.
Differences between male and female are not
limited to sexual organs and secondary sex characteristics; they are also
evident within the CNS. For example,
there is a sexually dimorphic nucleus of the preoptic area, which is
considerably larger in males and the same is true for a cell group in the
sacral spinal cord known as the nucleus bulbocavernosus.
The sex hormones, i.e. androgens and
estrogens, play important roles both in the development and differentiation of
the male and female sex organs and sexual behavior. GnRH producing neurons, responding to sensory
input and to circulating gonadal steroids, control the secretion of LH and FSH
from the anterior pituitary. GnRH secreting
neurons with projections to the portal system in the median eminence are
located in preoptic-anterior periventricular area. LH and FSH are released from the pituitary
into the systemic circulation in response to GnRH, and they travel to the
gonads, where they direct gamete production, as well as gonadal (testosterone
in male and estrogen and progesterone in female) hormone production.
Sexual behavior involves a number of general (e.g. respiratory and cardiovascular)
and specific (e.g. erection, ejaculation, etc) responses mediated in large part
by the autonomic nervous system.
Although several of these specific responses represent involuntary or
reflex phenomena, descending pathways from the hypothalamus or basal forebrain
regions play a significant modulatory role.
A critical brain area in male copulatory behavior seems to be the medial
preoptic area, whereas feminine sexual behavior appears to be more dependent on
regions in and around the ventromedial nucleus.
GnRH
neurons are born outside the brain in the olfactory placod and migrate
caudally to their final positions in the septal, preoptic and anterior
hypothalamic areas. GnRH release into the portal bloodstream occurs in a
coordinated fashion, with distinct pulses of GnRH secretion. The pulsatile
stimulation of the anterior pituitary by GnRH leads, in turn, to pulsatile
release of LH and FSH from the pituitary gonadotropes into the peripheral
bloodstream. Multiple-unit recording electrodes placed in the medial basal
hypothalamus of rhesus monkeys have measured spikes of electrical activity that
correspond in time to pulses of LH release. These bursts of electrical activity
may come from GnRH neurons themselves or from neurons that impinge upon the
GnRH neural system and thereby govern its firing pattern. The question of how
GnRH neurons, distributed diffusely throughout the hypothalamus, coordinate the
release of discrete pulses of GnRH into portal bloodstream remains unanswered.
GnRH neurons might actually form an interconnected network. Anatomical studies
showing synapses between GnRH neurons, and perhaps cytoplasmic bridges between
adjacent GnRH neurons.
Synaptic
input from a variety of neuronal types has been reported, including other GnRH
neurons; dopaminergic, noradrenergic, serotoninergic; neurons containing GABA,
CRF, substance P (SP), neurotensin (NT), Beta-endrorphin (B-END), etc.
GnRH
release is affected by the negative feedback of steroid hormones at
the level of the hypothalamus. Steroid hormone (estrogen, progesterone)
negative feedback decreases the frequency of pulsatile GnRH stimulation of the
pituitary and thus results in decreased frequency of pulsatile LH release (e.g.
luteal phase). At the level of the hypothalamus, steroid hormones likely
modulate the firing of neurons that project to LHRH neurons, since the LHRH
neurons themselves lack estrogen receptors (Figure 39).
Pulsatile
stimulation of the pituitary by GnRH is necessary to maintain normal function
of pituitary gonadotropes. There is a fairly narrow window of acceptable
frequency for stimulation of the pituitary by GnRH to achieve normal
gonadotropin secretion. Pulse frequency faster or lower than once per hour
usually leads to an inhibition of gonadotropin secretion. The gonadal
steroid hormones estradiol and testosterone can negatively feedback at the
levels of the hypothalamus and pituitary to decrease the frequency of pulsatile
gonadotropin secretion and decrease the amplitude of LH pulses. In women,
pulsatile LH secretion in the early follicular phase of the menstrual
cycle, when circulating concentrations of ovarian steroid hormones are quite
low, occurs at a frequency of approximately one pulse per hour. As the
follicular phase progresses, and the developing ovarian follicles begin to
secrete increasing amounts of estradiol, slows to one pulse every 90 min. In
the luteal phase of the menstrual cycle, when the ovary is secreting
large quantities of progesterone, as well as estradiol, LH pulses occur at a
frequency of once every 6-12 h. In contrast, in man, LH pulse frequency remains
rather stable throughout adulthood, at approximately one pulse every 2-3 hr.
However, testosterone levels play a large part in determining this frequency,
as seen by the effects of castration. Negative feedback of steroid
hormones at the level of pituitary decreases the sensitivity of pituitary
gonadotropes to GnRH and thus results in a decrease in the amplitude of LH
pulses but not the frequency of LH pulses.
Ovarian
function is cyclic (Figure 39).
In women, the first half of the menstrual cycle is characterized by the growth
and development of a cohort of follicles, culminating with selection and
ovulation of the ovum from a single dominant follicle at about 14 days from the
beginning of the cycle. After ovulation, the cells of the collapsed follicle
are reorganized into a transiently functional, steroid-producing gland, the
corpus luteum, that secretes progesterone and estrogen. Should fertilization of
the ovulated ovum occur, these ovarian hormones play a critical role in
preparing the uterus for implantation of the developing embryo. If a pregnancy
does not occur, the corpus luteum spontaneously becomes dysfunctional after
approximately 14 days. Withdrawal of gonadal hormones to the uterus leads to
menses and the period of follicular development is reinitiated. In women, and
other primates with menstrual bleeding, Day 1 of the menstrual cycle is
designated the first day of menstrual bleeding. Species, in which the
endometrium is reabsorbed, rather than sloughed, show a behavioral
predisposition to mating during the time of ovulation. Cycles in these species
are referred to as estrus cycle.
Estrogen and cognition. It is well established
that estrogens affect the brain throughout the life span. Moreover, the effects
are not limited to the areas primarily involved in reproduction but also
include areas relevant to memory, such as the basal forebrain and hippocampus.
For example, there is extensive evidence that estrogen levels are correlated
positively with dendritic spine densities within CA1 of the hippocampus and
that estrogen administration ameliorates learning deficits and cholinergic
abnormalities in ovariectomized rats (McEwen et al., 1997). Also, there are sex
differences in the rate of development and the magnitude of age related
impairments in spatial reference memory (Markowska, 1999) that is paralleled
with altered estrogen levels. There are several mechanisms for estrogen
actions: 1) estrogen may cooperate with nerve growth factor (NGF). For example
both estrogen and NGF receptors are expressed in the same basal forebrain
neurons and estrogen and NGF mutually enhances the binding of each other to its
receptors. 2) Estrogen induced dendritic changes are related to increased
expression of NMDA receptors and estrogen also enhanced LTP. 3) Estrogen may
act as antioxidant disrupting free radicals and protect beta-amyloid exposure
induced cell death. These findings may
explain the mild beneficial effect of estrogen-treatment in Alzheimer’s
disease.
The
HPA axis is a key player in an animal’s response to stressful stimuli. Other
participants are the adrenal medulla, which produces noradrenaline and
adrenaline and the sympathetic nervous system, which modulates physiologic
functions through neurotransmitters.
Corticotropin-releasing
hormone (CRH or CRF) is a 41 amino-acid peptide expressed in the
hypothalamus. The region with highest expression is the medial parvicellular
part of the PVN. CRH neurons in the mpPVN project to the external median eminence,
where peptides are secreted into the portal bloodstream, through which they are
transported to the anterior pituitary. In addition to CRH, the same neurons in
the PVN also express and release vasopressin, although most of the VP is
expressed in neighboring magnocellular elements of the PVN that project to the
posterior pituitary. Corticotropes in the anterior pituitary express receptors
for CRH and VP. In response to
stimulation, corticoctropes synthesize and release adrenocorticotropic hormone
(ACTH). ACTH through the systemic circulation binds and activates its receptors
on the surface of cells of the adrenal cortex. In response to receptor
activation, adrenocortical cells synthesize glucocorticoids.
Concentration of circulating
glucocorticoids and ACTH show a circadian rhythm. In humans, glucocorticoid
levels peak around 7 AM and decline steadily throughout the day. The nadir is
reached in the late evening at
Basal
activity of the brain-pituitary-adrenal axis oscillates: CRH is released in
a pulsatile manner from terminals in the
median eminence, but the system is activated under emergency conditions through
neural input. In the hypothalamus, the PVN appears to sum and integrate input
from numerous loci. Input to PVN is divided into several broad classes, like
brainstem (catecholaminergic fibers via the NTS convey viscerosensory
information); hypothalamic or limbic inputs (amygdala, septum, hippocampus,
prefrontal cortex reach PVN primarily via the bed nucleus of the stria
terminalis). Blood-borne signals through neural projection from SFO and OVLT
apparently also reach stress-related PVN parvicellular neurons.
Glucocorticoid
receptors are members of a superfamily of receptors that act as
ligand-regulated transacting receptors. In each case, the receptor protein
resides in the cytoplasm in a complex containing heat-shock proteins, which
fold the receptor into the appropriate configuration for recognizing
corticosteroid ligands. Upon steroid binding, the receptor moves to the nucleus
of the cell and interacts with specific hormone recognition (or response)
elements on the DNA, thereby changing transcription rate.
CRH
neurons of the PVN contain steroid receptors, and glucocorticoids inhibit
transcription of CRH and VP genes through genomic feedback however, numerous
other brain regions also express steroid receptors. These regions, including
the hippocampus exert negative feedback on the brain-pituitary-adrenal axis
through projections to PVN.
The
negative feedback loop of CRH-ACTH-glucocorticoids is kept in a delicate
balance. In response to stress, there is a large increase in the activity of
the stress axis, but the system is downregulated rapidly by negative feedback
from the glucocorticoids to the brain and pituitary gland, causing it to return
the output of the stress axis to basal levels. However, the stress axis can be
disrupted by psychological stressors such as mood disorders that can
chronically disregulate the HPA axis (PTSD). Hyperadrenocorticism (Cushing
syndrome) is concomitant with immunosupression, osteoporosis, and muscle
atrophy. Underactivity of the stress axis (
9. Central control of food intake (Figures 46,
47, 48, 49, 50)
Food-intake is a complex process in which various hypothalamic neurons (PVN, arcuate, ventromedial and lateral hypothalamic–perifornical neurons (LHA, PFA) are participating that integrate sensory inputs from the viscera and influence autonomic outflow to viscera. Similarly a host of transmitters (noradrenaline, serotonin, dopamine) peptides (NPY, CART) and blood- borne substances (insulin, leptin) with their receptors on various hypothalamic neurons are involved in the central regulation of food intake.
According to a recent integrated model (Figures 46-47), the adiposity signals, leptin and insulin stimulate a catabolic pathway via POMC/CART neurons and inhibit an anabolic pathway (NPY/AGRP) both originating in the arcuate nucleus. These pathways project to the PVN and LHA/PFA, where they make connections with central autonomic pathways that project to brainstem autonomic regions that process satiety signals. Afferent input related to satiety from the gastrointestinal tract (gastric distension), from gut peptides such as CCK (which is secreted during meals) are transmitted through the vagus nerve to the nucleus of the solitary tract (NTS), where they are integrated with descending hypothalamic input from leptin/insulin sensitive neurons. Also, the NTS is the site where sensory input from the viscera is integrated with input from the taste buds. Net neuronal output from the NTS leads to termination of individual meals, and is potentiated by catabolic projections from the PVN and inhibited by input by anabolic input from LHA/PFA.
Arcuate
nucleus (Figure 48). The
highest density of leptin receptor mRNA is in the ventrobasal hypothalamus,
including the arcuate nucleus. Microinjection of leptin into the arcuate
nucleus area results in anorexic response. Leptin signals in the arcuate
nucleus are mediated by POMC (pro-opiomelanocortin: alpha MSH) and CART
(cocaine- and amphetamine-regulated transcript)-containing efferents that are
activated by the action of leptin.
Alpha-MSH and CART are potent catabolic peptides, and when either is
administered locally in the 3rd ventricle, animals eat less food,
have increased energy expenditure, and lose weight. The other type of arcuate
neuron influenced by adiposity signals synthesizes neuropeptdie Y (NPY) and
agouti-related protein (AGRP). Both
peptides are potent anabolic compounds in that the administration of either
into the 3rd ventricle results in hyperphagia, reduced energy
expenditure and weight gain. Neurons containing NPY and AGRP express both
leptin and insulin receptors and the local administration of either insulin or
leptin near the arcuate nucleus reduces the synthesis of both NPY and AGRP.
Expression of NPY mRNA in the arcuate nucleus is elevated in response to
fasting and in leptin-deficient ob/ob and leptin-resistent db/db mice. In ob/ob
mice and fasted rats, exogeneous treatment with leptin suppresses NPY
overexpression. Endogeneous levels of
NPY in the arcuate-PVN system normally peak when daylight ends and nocturnal
activity begins, which is also the time when rats typically eat their largest
meal of the day.
Paraventricular
nucleus (Figure 48). The PVN is uniquely equipped to control
activities both in the endocrine, autonomic and somatomotor systems. It is populated by both magno and
parvocellular neurosecretory neurons, that project to the neurphypophysis and
to the portal system in the median eminence, thus can affect thyroide hormone,
growth hormone and ACTH secretion. The
regulation of the hypothalamo-pituitary adrenal axis, which is controlled by CRF
secreting neurons, may be of special importance in this context. For instance, corticosterone affects the
carbohydrate metabolism, and it is well known that different forms of stress
can influence eating behavior. Parvocellular neurons in the PVN also project to
the brainstem parasympathetic and to the sympathetic preganglionic autonomic
nuclei in the medulla and spinal cord.
Through these pathways, the PVN can directly influence the hormone
secretion from pancreas and adrenal medulla as well as somatomotor activities,
that may be relevant in feeding behavior. Leptin treatment activates neurons in
the PVN. The PVN may regulate many of the responses of leptin, it receive
projections from leptin-responsive neurons of the arcuate nucleus and has
chemically and anatomically specific projection to brain control sites involved
in the maintenance of autonomic and endocrine homeostasis. Oxytocin secreted from PVN neurons
but projecting within the CNS rather than to the pituitary has also appetite
suppressive effect.
Figure 48 show the localization of NPY and POMC (alpha MSH) neurons in the arcuate nucleus and their projections to oxytocin, CRH and TRH neurons in the PVN (which cause anorexia). Arcuate POMC axons also contact orexin and MCH neurons in the LHA/PFA areas whose action increase feeding. Figure 48 shows the putative mechanism of obesity and anorexia in leptin/insulin deficiency and increased leptin/insulin signaling, respectively. Reduced leptin/insulin levels in the brain during diet-induced weight loss increases activity of anabolic pathways that stimulate eating. Increased leptin level by inhibiting NPY and facilitating alpha-MSH neuronal systems results in anorexia.
Ventromedial hypothalamic nucleus (VMH). Although recent work does not support the
conclusion from early lesions studies that VMH functions as a satiety center,
VMH neurons are rich in insulin and leptin receptors that mediate adiposity
signals.
Lateral hypothalamic (LHA) and Perifornical (PFA) areas (Figure 48). Lesions of the
lateral hypothalamus (LHA) can cause decreased food intake. These animals show akinesia and sensory
neglect. In this respect, rats with lateral hypothalamic lesions resemble human
patients with Parkinson’s disease that has been attributed to the degeneration
of dopaminergic neurons of the nigrostriatal system. Large lesions in the LHA
interrupt the ascending dopaminergic fibers from the ventral midbrain.
Similarly, selective damage to the dopaminergic neurons with 6-OHDA also produces
akinesia, sensory neglect in association with loss of food intake. Neurons in
the LHA, including those that contain melanin-concentrating hormone
(MCH) project to the cerebral cortex. Ob/ob (leptin deficient) mice have
elevated level of MCH mRNA and this overexpression is normalized by leptin
administration. Another peptide, orexin/hypocretin
is found in the perifornical region projecting to several areas of the
neuraxis, including the cerebral cortex may be important in mediating leptin
effect. Administration of MCH or orexin into the brain stimulates food
intake.
Leptin
is a circulating hormone produced by white adipose tissue and has potent
effects on feeding behavior, thermogenesis and neuroendocrine response. Leptin
regulates energy homeostasis, its absence in humans and rodents cause severe
obesity. Leptin concentration in plasma is directly proportional to adiposity.
Congenital leptin deficiency resulting from mutations within the leptin gene
causes extreme obesity (ob/ob mice). Moreover, a mutation in the leptin
receptor gene (db/db mice) results in morbid obesity, failure to undergo
puberty and decreased level of growth hormone and thyroid hormone. However,
most obese humans do not have mutations in the leptin or leptin-receptor genes
but have high levels of circulating leptin that fail to prevent obesity. By
analogy with diabetes mellitus, these people may have functional leptin
resistance and impaired responsiveness to circulating leptins. Administering
leptin to ob/ob mice, or to normal animals, causes them to eat less and loose
weight. Hence, leptin appears to function as a negative feedback signal to the
brain. When fat stores increase in adipose tissue, more leptin is secreted and
enters the brain, causing a greater inhibition of food intake and loss of body
fat. When circulating leptin levels are low, feeding and other anabolic
responses are disinhibited. In this way, leptin acts to promote the maintenance
of a relatively stable body weight over long intervals.
Within the brain, leptin’s main site of action seems to be the hypothalamus. Figure 49 shows that the effect of leptin is mediated through leptin receptors localized in several hypothalamic nuclei, including, the ventromedial, (VMH) arcuate and dorsomedial nuclei. The complex physiological effect of leptin can be explained by the fact that leptin-responsive neurons in these regions project to hypothalamic neurons in PVH, LHA and subparaventricular zone. These neurons in turn with their further projections to the cerebral cortex, autonomic preganglionic neurons and the pituitary gland can affect various endocrine, autonomic and behavioral responses.
Leptin
acts directly through hypothalamic long form leptin receptors. The leptin receptor have a docking site for janus
kinases (JAK), a family of tyrosine kinases involved in intracellular cytokine
signaling. Activated JAK phosphorylates members of the signal transduction and
transcription (STAT) family of
intracellular proteins. STAT proteins, in turn, stimulate transcription of
target genes that mediate some of leptin’s cellular effect. Leptin can affect
neuronal firing rate independently of its transcriptional effect. This response is absent in db/db mice, which
lack the functional long-form leptin receptor, and thus cannot activate STAT
proteins.
Leptin
also activate immediate early genes in specific nuclear groups. Intraveneous
leptin activates several regions thought to be involved in regulation of energy
balance, including VMH, DMH and PVN nuclei. Leptin administration also
activates cells in the superior-lateral parabrachial nucleus that project a CCK
containing pathway to the VMH. Recently
a leptin-inducible inhibitor of leptin signal transduction has been described,
which is rapidly induced in the hypothalamus following systemic leptin
administration.
Noradrenaline (NA) is synthesized in brainstem locus coeruleus and NTS region (A1
noradrenergic cell group). These areas project both caudally to the spinal cord
and rostrally to the hypothalamus, thalamus and cortex. In some of these
neurons, including those projecting to the PVN, noradrenaline is colocalized
with NPY. Like NPY, injection of noradrenaline into the PVN increases food
intake. The observation of elevated NA levels in the PVN of ob/ob mice may
indicate that increased NA signaling in the PVN may contribute to hyperphagia
induced by leptin deficiency.
Dopamine.
Feeding effects of dopamine vary with the brain region under study. For
example, mesolimbic dopamine pathways, originating from the SN-VTA that project
to the nucleus accumbens, striatum and cerebral cortex seem to contribute to
the rewarding aspects of palatable food. In contrast, arcuate DA neurons seem
to inhibit food intake. In ob/ob mice the reduced arcuate DA level may
contribute to hyperphagia induced by leptin deficiency.
Serotonin.
Several centrally acting drugs developed for obesity treatment (e.g.
dexflenfluramine) increase 5HT receptor signaling and thereby suppress food
intake, whereas antagonists have the opposite effect. The 5HT2C
subtype is implicated in this process as knockout of this receptor increases
food intake. Leptin increases 5HT turnover and thus it is possible that at
least some of leptin’s weight-reducing effects are mediated through increased
5HT signaling.
The
circadian timing system is composed of central and peripheral neural elements.
Critical components include the photoreceptors and visual pathways (e.g
retinohypothalamic tract to the SCN), circadian clocks or pacemakers (such as
the SCN) and output pathways (pineal gland, melatonin) to couple pacemakers to
effectors (Figure 51E).
SCN neurons are spontaneously active circadian oscillator even when deprived of
the afferent signals. With SCN lesions,
the sleep-wake rhythm is eliminated, but the amount of time spent asleep and
awake and the amount of REM and nonREM sleep is unaffected. Ablation of the SCN
results in also a loss of rest-activity rhythm, estrus cycle and reproductive
capacity in rats (Figure 52C). It is also
clear the SCN influences the temporal organization of feeding. For example,
nocturnal rodents normally feed almost exclusively during the dark phase of the
photoperiod, but rodents without SCN feed throughout the light-dark cycle.
Genetically controlled molecular clock (Figures 52a,b). Individual SCN neurons maintained in cell culture
have a rhythmic electrical activity that approximates a 24 h cycle.
Interestingly, SCN cells show increased electrical activity during the daytime
in both nocturnal and diurnal mammals, thus SCN activity signals inactivity in
nocturnal animals (rats, mice) and activity in diurnal species such as humans.
In humans, however, secretion of corticosteron (Figure 55) take place opposite
to that observed in the rat, so the signal of the SCN to the human PVN and
other hypothalamic targets will be interpreted in a different way.
Mammalian
circadian rhythms are maintained intracellularly by interlocking positive and
negative-feedback control of the transcription and subsequent translations to
protein of about a dozen of clock genes that reliably recur at precise times
over 24-h cycles. These molecular signals can be read by cytoplasmic mechanisms
in SCN cells and translated into cellular events such as changes in membrane
potential and cell firing rate. Such signals, in turn, can be transmitted to
connecting neurons and ultimately to those neuronal structures that control
physiological processes with a circadian rhythmicity. Evidence suggests that
GABA, gap junctions and neural cell adhesion molecules participate in the
coupling of individual SCN neurons that underlie pacemaker function.
Synchronization of the biological clock with the light-dark cycle (entrainment). The circadian rhythms of physiology and behavior are driven by the molecular clock as indicated above. To be ‘useful’, these clocks must be synchronized to the daylight cycles of the real world. The primary environmental synchronizing cue is the natural cycle of light and dark. In fish, amphibians, reptiles and birds there are specialized circadian photoreceptor cells located in several places in the brain. These cells respond to light that penetrates the skin, skull and overlying brain tissue, and their output signals act directly on clock centers in the brain. In these animals, eyes are not necessary for synchronization to the day-night cycle. Mammals have specialized photoreceptors in the retina. Light establishes both the phase and period of the pacemaker, and thus is the dominant entraining stimulus or Zeitgeber (time-giver) of the circadian system. The pacemaker can be viewed as a somewhat inaccurate clock, which must be repeatedly reset. It free runs with a period that is slightly off 24 h in the absence of light-dark cycle. The light-dark cycle sets the exact timing of the pacemaker and it is best understood by looking at the phase response curve (PRC) of the pacemaker to light (Figure 54A). The PRC shows that the pacemaker responds differently to light at different times of the day. It is typically reset each day in the morning and the evening at the transitions between light and dark.
The
circadian system responds to changes in luminance, the total amount of light,
but not to color, shape, movement or other visual parameters. The responsiveness of the circadian system to
light is not altered in mutant mice with retinal degeneration and nearly
complete loss of rods or cones. Recently, it has been shown that a small
percentage (1-2% in the rodent’s retina) of retinal ganglion cells that project
to the SCN and the lateral geniculate body (specifically to the intergeniculate
leaflet (IGL) are intrinsically light-sensitive, contain a photopigment
melanopsin (an opsin based photopigment), and register luminance (Hattar et
al., 2002; Berson et al., 2002) (Figure
53). These same cells also contain the peptide PACAP (pituitary
adenyl-cyclase-activating peptide). These cells respond electrically to light
in isolated retinal preparation in which synaptic transmission is blocked. It
is suggested that this, separate visual circuit, running in parallel with the
image-forming visual system, encodes the general level of environmental
illumination and drives synchronization of the biological clock with the
light-dark cycle. This ‘new’ light-detection system influences also to pupil’s
response to light and suppression of melatonin secretion produced by light.
Melanopsin-deficient mice could still be entrained to a L/D cycle, still
exhibited phase-shifting in response to pulses of white light and responded
with changes in circadian period when they were switched from constant darkness
to constant light, but these responses were severely attenuated, indicating the
critical role of melanopsin in circadian photo entrainment. These
melanopsin-deficient animals had also a diminished pupillary light reflex at
high irradiance. It is suggested that the full dynamic range of the pupillary
response and other components of the circadian mechanisms could be accounted
for by the rode/cone, melanopsin and cryptochrome (another photopigment present
in the inner retina and SCN) systems acting together (Figure 54B).
The
IGL modulates entrainment by transmitting information about both photic and
nonphotic (locomotor) events to the SCN.
The retinal ganglion cell-SCN circuit uses glutamate as transmitter, the
IGL uses GABA and NPY. Perfusion of NPY into the SCN or IGL stimulation
produces phase response curve with phase advances during subjective day and
phase delays during subjective night. Serotoninergic neurons of the midbrain
raphe densely innervate SCN neurons. Raphe neurons are state-dependent; they
fire regularly during waking, slowly during SWS, and not at all during REM
sleep. During waking visual stimulation acutely increases the activity of SCN
neurons. Serotonin inhibits the SCN response to light. Data suggest that
serotonin acts between the retina and pacemaker mechanisms within SCN neurons.
Specific receptors and signaling
pathways in entrainment. The rhythm of SCN cell firing depends on specific membrane receptors,
signaling pathways and the timing of various neuromodulators as they relate to
circadian phase. Photic and glutamatergic stimulation of SCN cells in
the early subjective night cause phase delay, that is mediated by ryanodine
receptors and elevated intracellular calcium, whereas such stimulation late in
the subjective night causes phase advance that is mediated by a cyclic GMP
pathway. By contrast, photic resetting of the circadian clock during the
subjective day is believed to be mediated by pituitary
adenyl-cyclase-activating peptide (PACAP) and cyclic AMP. Cholinergic mechanisms that involve the
activation of M1 muscarinic receptors in the SCN are involved in resetting the
circadian clock only during the subjective night. Similarly, SCN cells are most
sensitive to melatonin feedback at subjective dawn. Second-messenger cascades
lead ultimately to the intranuclear phosphorylation of
cAMP-responsive-element-binding protein (CREB) and the subsequent downstream
transcription of clock-related genes.
Output of SCN neurons controls various rhythms The SCN project to the subparaventricular zone
(SPVZ) and other hypothalamic neurons and several other nonhypothalamic
structures of the diencephalons (Figure
55). Direct activation of these efferent circuits controls the
expression of circadian rhythmicity. SCN projections have peak daytime firing
rates that are about twice as fast as night rates. However, transplantation
studies also suggest that in addition to synaptic humoral mechanism may play a
role in rhythm regulation. The SCN uses
several means to regulate circadian rhythm of hormonal secretions: 1) by direct
contact with neuroendocrine neurons, for example those containing GnRH and CRH;
2) by contacting neuroendocrine neurons via intermediate neurons, for example
those of the medial preoptic area and dorsomedial hypothalamic nucleus and 3)
by projections to the autonomic PVN to influence the autonomic nervous system
(for example in the case of melatonin regulation)
Estrous cycle In female mammals, reproductive events occur in cycles, called
estrous cycles. The estrous cycle in the rat is four days long and culminates
on the day of proesrtrus in a mid-afternoon surge of release of LH. The timing
of the LH surge is precise and results in ovulation followed by receptivity to
males the subsequent night. The surge in LH production results from GnRH
release from the median eminence into the portal circulation. The precise
timing of the release of GnRH to produce the LH surge is a function of a series
of endocrine and neural events that lead to proestrus, with the SCN providing
the crucial temporal signal. SCN ablation results in loss of estrous cycles and
reproductive capacity.
Melatonin regulation (figure 56). In habitats with marked seasonal variation of temperature
and food supply, survival of species requires seasonal regulation of
reproduction. The circadian timing system of these animals uses production of
the pineal hormone melatonin to measure day length as a means of predicting
seasonal changes. The SCN via the PVN-intermediolateral column-superiocr
cervical ganglion (SCG)–pineal gland routes control melatonin secretion that
faithfully mirrors the lengths of day and night. For example, in all mammals,
the activity of SCG neurons innervating the pineal increases at night. The
release of norepinephrine from axon terminals acts through beta-adrenergic
receptors to stimulate melatonin synthesis and release. At night, exposure to
light through the retinohypothalamic tract via GABAergic projection from SCN to
PVN inhibits SCG activity and thereby quickly stops the production of
melatonin. In the mammalian brain,
melatonin receptors were identified in the SCN, the midline thalamic nuclei and
the pars tuberalis of the anterior pituitary. Melatonin can affect pacemaker
function via its action on SCN cells. For example, melatonin can lessen the
symptoms of jet lag and in elderly people, the disruption of sleep as a
consequence of alteration of the pacemaker function.
Sleep-wake cycle (Figures 57-58). The function of the circadian timing system is
to coordinate a large series of humoral, physiological and behavioral
mechanisms to promote maximally effective sleep and adaptive waking behavior.
In a series of elegant experiments, Aston-Jones et al. (2001) have shown that
an SCN-Dorso Medial Hypothalamic (DMH)-locus coeruleus (LC) pathway is
necessary for circadian rythmicity in the LC firing. The transmitter of the
DMH-LC may be orexin/hypocretin. This route may be important in circadian
regulation of sleep and waking. It is
also suggested that the SCN provides arousal-promoting input to hypocretin
neurons, which project upon the neocortex and subcortical arousal areas (
The
hypothalamus plays an important role in sleep regulation. Lesion of the
preoptic region produces insomnia in rats, whereas chemical or electrical
stimulation of this region causes sleep. In contrast, lesions of the caudal
hypothalamus produce somnolence, suggesting that this region is involved in
arousal. Recently, Saper and colleagues (Sherin et al; 1996), using the
localization of the protein product of the immediate-early gene c-fos, defined
the location of hypothalamic neurons that are active during sleep. These,
(presumably GABAergic) cells, located in the ventrolateral preoptic region
(VLPO), project to the caudal hypothalamic histaminergic cell groups
in the tuberomammilary nucleus (TMN), which diffusely innervate the cerebral
cortex and promotes arousal.
Several cell groups in the hypothalamus (see SUBCORTICAL MODULATORY SYSTEMS) project diffusely to the cerebral cortex and have been postulated to be important in arousal. Recently, a 130 amino acid containing protein, called hypocretin (orexin) was isolated from the hypothalamus (Sutcliffe et al, 1997). In situ hybridization and immunocytochemical studies revealed that neurons expressing this protein are located exclusively in the tuberal region of the hypothalamus around the fornix. Although it is a restricted group of cells, their projections were widely distributed in the brain, including a diffuse projection to the cerebral cortex. Knockout mice that lack the gene encoding this protein show narcolepsy, indicating that this protein is important in arousal.
Saper suggests that between sleep and wake-promoting brain regions reciprocal interactions exist which results in stable wakefulness and sleep (Fig. 31). Disruption of wake- or sleep-promoting pathways results in behavioral state instability. For example, after lesion of the VLPO, the animals experience much more wakefulness. Similarly, hypocertin-knockout mice or narcoleptic persons fall asleep more rapidly than unaffected individuals.
12. Central control of osmo- volume
regulation. Thirst. Drinking
Body fluid homeostasis is directed at achieving
stability in the osmolality of body fluids and the volume of the plasma. Such
homeostatic regulation is promoted by several mechanisms intrinsic to the
physiology of body fluids (intar-extracellular) and the cardiovascular system.
For example, the osmotic movement of water across cellular membranes rapidly
buffers changes in the osmolality of extracellular fluid. Similarly, the
movement of fluid across capillary membranes buffers acute changes in plasma
volume, as does venous compliance and compensatory alterations in the kidney
glomerular filtration rate. Nevertheless, changes in body fluid osmolality and
plasma volume maybe so large that additional mechanisms must be recruited to
maintain homeostasis. These responses include central control of water and
sodium excretion in urine through specific actions of hormones, and the central
control of water and sodium consumption motivated by thirst and salt appetite
(Striker and Verbalis, 2002).
Fluid homeostasis is regulated by several
interdependent mechanisms, one of which, i.e. the retention of water by
the kidney mediated by the hypothalamo-hypophysial vasopressin system, was
discussed above. Increased water
intake is another mechanism of replenishing body fluids. Although we often drink spontaneously,
drinking can also be activated by water deficit, i.e. deprivation induced
drinking. Deprivation-induced drinking
is regulated primarily by osmotic changes in the blood or a change in blood
volume, e.g. hemorrhage. The
osmoreceptors for drinking, like the ones regulating vasopressin release, are
located in the OVLT and neighboring medial preoptic area, near the anterior wall
of the 3rd ventricle (AV3V).
Drinking in response to reduced blood volume is initiated by two
different mechanisms. One type of input
originates in mechanoreceptors in the pulmonary artery and the vena cava, and
reaches the hypothalamic integration centers for drinking via the nucleus of
the solitary tract. Another important
stimulus is blood-borne, when blood pressure falls, the kidneys release renin
into the bloodstream. Renin triggers a biochemical cascade that produces
angiotensin II, which activates the neural circuit for drinking behavior
through its action on the subfornical organ.
As indicated above, the subfornical organ is also an intermediary
structure in another important mechanism for fluid homeostasis, i.e. the place
through which blood-borne angiotensin II (which is released in increased amount
in response to reduced blood volume) can activate the vasopressin system to
reduce the loss of water through the kidney. The neural pathways connecting the
SFO and AV3V with magnocellular cells in the SON and PVN have been identified;
the neural circuits in the forebrain that control thirst are still unknown.
When body fluid is hyperosmolal, adaptive behavior includes not only drinking and conserving water, but also excreting sodium and avoiding the consumption of additional osmoles. Endogeneous natriuretic agents promote urinary sodium loss after an administered sodium load. One such agent is the hormone atrial natriuretic peptide (ANP), which is synthesized in the atria of the heart and released when increased intravascular volume distends the atria. ANP is also synthesized in central neurons of the hypothalamus. Another is the hormone oxytocin. Like VP, OT is secreted from the posterior pituitary in proportion to induced hyperosmolality. OT is as potent in stimulating natriuresis as VP is in stimulation antidiuresis. Renal Na+ retention is mediated largely by aldosterone secreted from the adrenal cortex. The secretion of aldosteron is stimulated by ANGII and ACTH. Note that aldosterone can eliminate Na: from urine, whereas VP primary effect is antidiuresis. Circulating oxytocin also inhibit salt appetite.
13. Temperature Regulation
In order to sense changes in body temperature and in
the surrounding temperature, thermoreceptors are located largely in the skin
(free nerve endings) and in the brain, where thermosensitive neurons are found
primarily in the preoptic-anterior hypothalamic area (PO-AH). The thermosensitive neurons in the PO-AH
sense the temperature of the blood that passes through this richly vascularized
region. Based on this information and
that from various peripheral receptors, neural assemblies both in the anterior
and posterior hypothalamus coordinate the activity needed to maintain the body
temperature within fairly narrow limits.
In response to increased body temperature, hypothalamic neurons initiate
a series of processes that result in heat -loss, including peripheral vasodilatation
and sweating. A drop in the surrounding
temperature leads to a series of events including peripheral vasoconstriction,
piloerection, increased metabolism and shivering in order to preserve heat.
Thermosensitive neurons appear to influence arousal mechanisms integral to
regulation of the sleep-wake cycle. For example, Dennis McGinty and colleagues
have shown that warming the preoptic region during waking suppresses
arousal–related neuronal activity in the caudal hypothalamus and in
magnocellular basal telencephalon neurons that have diffuse cortical
projections. The consequence of this experimental warming includes decreased
motor activity, reduced metabolic activity and respiratory rate and enhanced
peripheral heat loss. All these effects are similar to changes that
characterize the onset of sleep.
The preoptic region of the hypothalamus also plays a role in producing fever. Circulating cytokines may act at or near the OVLT to elicit fever, and prostaglandin injection induces fever has been identified in the region surrounding the OVLT.
Aston-Jones et al: A neural circuit for circadian regulation
of arousal. Nature Neuroscience 4:
732, 2001.
Buijs and Kalsbeek: Hypothalamic integration of central and peripheral
clocks. Nature Reviews Neuroscience 2:521, 2001
Card et al: The Hypothalamus: An overview of regulatory
systems. In: Fundamental Neuroscience (Squire et al., eds), 2nd
edition, p. 897, 2002
Gore and Roberts: Neuroendocrine Systems. In: Fundamental
Neuroscience, p. 1031.
Elmquist et al. Unraveling the central nervous system pathways
underlying responses to leptin. Nature Neuroscience 1:445, 1998.
Hattar et al: Melanopsin-containing retinal ganglion cells:
architecture, projections, and intrinsic photosensitivity. Science
295:1065, 2002.
Markowska:
Sex dimorphism in the rate of age-related decline in spatial memory:
Relevance to alterations in the estrous cycle.
J. Neuroscience 19: 8122, 1999.
Menaker:
Circadian
photoreception. Science 299:213, 2003.
Saper
et al: The sleep switch: hypothalamic control of sleep and wakefulness. Trends
Neurosci, 24, 726, 2001.
Schwartz
et al: Central nervous system control of food intake. Nature 404, 661,
2000.
Striker and Verbalis: Water intake and body fluids. In: Fundamental Neuroscience, p.
1011
Woods and Stricker: Food
intake and metabolism. In: Fundamental
Neuroscience, p.
991
15. Legend to figures
Fig.1. Summary of autonomic and behavioral regulation of hypothalamic origin.
Fig.2. Summary of neuroendocrine regulations by the hypothalamus.
Fig.3 Schematic diagram of the target organs upon which pituitary hormones act (Kendell and Schwartz)
Fig.4. Median sagittal section of the human brain showing the hypothalamus
Fig.5 Close-up view of the hypothalamus. The grey substance around the fornix (F) and mammillothalamic tract (MT) is removed. OCH= optic chiasm; LT=lamina terminalis.
Fig.6. 3-D diagram to show the hypothalamic nuclei.
Fig.7. Base of the brain.
Figs.8-11. Coronal sections from the Jakoklew Collection. Dark areas myelinated fiber bundles, yellow: gray matter.
Figs.12-14. Coronal series of human brain sections stained for Nissl. (Saper, 1990). DB= nucleus of the diagonal band of Broca; SO=supraoptic nucleus; SCN=suprachiasmatic nucleus; PVH=paraventricular hypothalamic nucleus; OT=optic tract; MPO=medial preoptic area; SI= substantia innominata; AHA=anterior hypothalamic area’; ARH=arcuate nucleus; VMH=ventromedial hypothalamic nucleus; DMH=dorsomedial hypothalamic nucleus; LHA=lateral hypothalamic area; TM=tuberomammillary nucleus.
Fig. 15. Schematic
coronal section through the human hypothalamus (Le Gros Clark, from Heimer,
1995).
Figs.16-17. Coronal sections of the rat hypothalamus stained for Luxol fast blue-cresyl violet..
Fig. 18. Horizonatl section through the rat hypothalamus (from Palkovits-Zaborszky, 1979) showing the medial forebrain bundle (MFB).
Fig. 19. Horizontal (A), sagittal (B) section, block scheme (C) to show the location and connections (D) of the fornix,
Fig. 20. Scheme of the medial corticohypothalamic tract.
Fig.21. Sagittal sections to show the course of the mammilothalamic (FMT), fasciculus mammilaris princes (FMP), fornix (F), stria medullaris (SM) and fasciculus retroflexus (habenulo-interpeduncular tract). cm=corpus mammilare
Fig.22. A: Schematic 3D-drawing to show the course of the stria terminalis, B: ventral amygdalofugal pathway.
Fig.23. Brainstem (yellow=dopaminergic; blue=serotoninergic and red=adrenergic-noradrenergic) pathways to the hypothalamus;
Fig.24. intrahypothalamic connections
Fig.25. Efferent hypothalamic connections,
Fig.26. A: concept of neurosecretion: hypothalamic neuroendocrine cells release secretory products into the blood stream. B: Control of the anterior and posterior pituitary by the hypothalamus.
Fig.27. A: schematic sagittal section showing the basic design of the median eminence; B: semithin section through the median eminence. Upper lining ependyma of the 3rd ventricle. C: Localization of somatostatin containing cell bodies (in the periventricular and the arcuate nucleus) and their terminals in the median eminence (Reichlin). D: Schematic view of the median eminence in coronal section.
Fig.28. Electron micrograph showing the external zone of the median eminence with the terminals of the parvocellular neurosecretory axons and the fenestrated capillaries (arrow). Ps=perivascular space.
Fig.29. Schematic drawing depicting troph-hormone producing cells in the anterior pituitary. In the prolactin cell 1-7 depict the secretory process from the endoplasmatic reticulum (1) through Golgi (3) and exocytosis (7). 6a-6b= lysosomal degradation of the secretory product. Note fenestrated capillaries just like in the median eminence.
Fig.30. Subdivision of the paraventricular hypothalamic nucleus showing the separation of magnocellular (oxytocin, vasopressin) and parvocellular neurons (from Swanson and Sawchenko)
Fig. 31. Schematic darwing to show the major subdivisions, afferent and efferent connections of the PVN (Swanson and Sawchenko)
Fig. 32. A-B: Distribution of oxytocine and vasopressin-containing neurons in the PVN (A) and (B) SON. C: transection of the hypothalamo-hypophysial fibers results in proximal (left) and distal (right) accumulation of neurosecretory products.
Fig.33. A-B: synapses in the SON. Note in A the presence of large (2,000A) neurosecretory granula in axons. D=dendrite, Sp=spine, A=axon, Cb=Cell body. B: Number and proportion of synaptic terminals in the SON (Zaborszky et al., 1975).
Fig.34. Schematic diagram of the peripheral stimuli known to elicit coordinated and preferential release of oxytocin and vasopressin from hypothalamic magnocellular neurons, as well as the resultant effect of these hormones on distant target organs. (Sawchenko).
Fig.35. A schematic drawing of a sagittal section through the rat brain to summarize current understanding of pathways involved in the reflex control of vasopressin (AVP) and oxytocin (OT) secretion from the posterior pituitary (Sawchenko)
Fig.36. Schematic diagram of the possible neural substrates and pathways mediating the baroreceptor-induced inhibition of vasopressin secretion in the SON neurons in the rat. LC=locus coeruleus, NTS=nucleus tractus solitarius, A2= nordarnergic neurons within the NTS area; A1/C1= adrenergic-noadrenergic neurons in the ventrolateral medualla, CH=cholinergic.
Fig.37. Vasopressin can act at three different sites in the body to modify cardiovascular function. It can potentiate the vasoconstrictor action of noradrenaline at blood vessels, modulate sympathetic transmission, and affect the baroreceptor reflex by a central effect mediated by the area postrema.
Fig.38. The renin-angiotensin system. (Stricker and Verbalis, 2002)
Fig.39. Cyclic ovarian function, estrogen, LHRH, LH. A: normal menstrual cycle. Note changes in ovarian, uterine histology and peripheral hormone levels. B: Histological changes in the ovarium and uterus in case fertilization occurs. C: Changes in the hypothalamic concentration of LHRH and peripheral plasma concentration of LH throughout the proestrus in the rat. Note that preceding the release of the ovulation-inducing surge of LH there is a dramatic decline of hypothalamic concentration of LHRH and a corresponding increase in portal blood levels of LHRH. D: Concentration of LH, FSH, estradiol (E2) and progesterone (Prog) in serum samples taken daily throughout the menstrual cycle of nine woman. The data are normalized to the day of the LH peak.
Fig.40. Pulsatile LH release in human female and rats.
Fig.41. Neural and humoral regulation of LHRH neurons.
Fig.42. Schematic representation of the HPA axis. Distribution of CRF neurons in the rat brain.
Fig.43. Schematic representation of the central stress circuitry (Herman, Cullinan).
Fig.44. Simplified scheme to suggest possible mechanisms in stress leading to hyperadrenocorticism (courtesy of Dr. Cullinan)
Fig.45. Stress-induced hippocampal dendritic atrophy and its pharmacological prevention (McEwen and Magarinos, 1997).
Fig.46. Relation between adiposity and food intake (Schwartz et al., 2000).
Fig.47. Neuoanatomical model of pathways by which adiposity signals interact with central autonomic circuit regulating meal size (Schwartz et al., 2000)
Fig. 48. Hypothalamic neuropeptides that influence caloric homeostasis (Schwartz et al., 2000; Woods and Stricker, 2002).
Fig.49. Neuroanatomical model of leptin action in the brain. (Saper)
Fig.50. Role of arcuate nucleus neurons in adiposity signaling (Schwartz et al., 2000).
Fig.51. Overview of the basic organization of the
mammalian circadian system (
Fig.52. a-b: The molecular control of circadian
regulation (Pace-Schott and Hobson, 2002). c: in the absence of SCN the
circadian regulation of rest-clock activity is lost (
Fig. 53. Connection of retinal pigment-containing ganglion cells (Hattar et al., 2002; Moore, 2002)
Fig.54. Light entrainment with intact and without molecular clock (Moor, 2002; Buijs and Kalsbeek, 2001)
Fig. 55. Effect of circadian signal from the SCN on various hormonal effector systems (Moor, 2002; Buijs and Kalsbeek, 2001)
Fig.56. Circadian regulation of melatonin secretion (
Fig.57. Model for reciprocal interactions between sleep and wake promoting brain regions (Saper et al., 2001)
Fig. 58. Circadian regulation of sleep-wake cycle (Pace-Schott and Hobson, 2002, Moore, 2002)
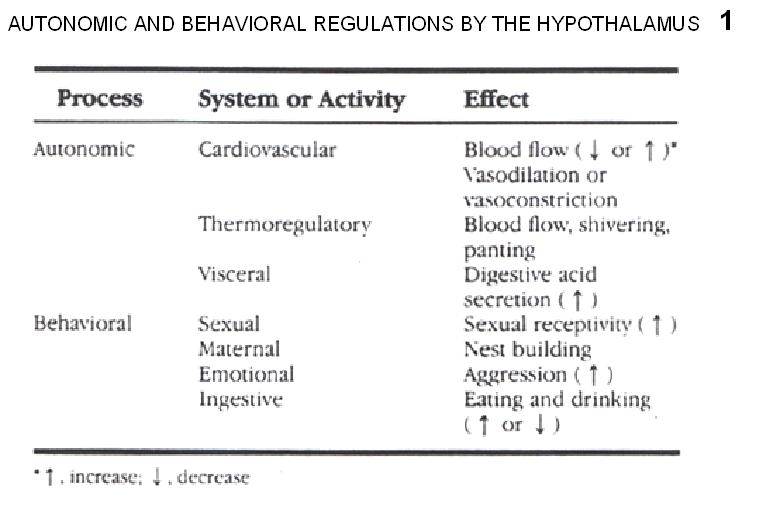{kind=link}
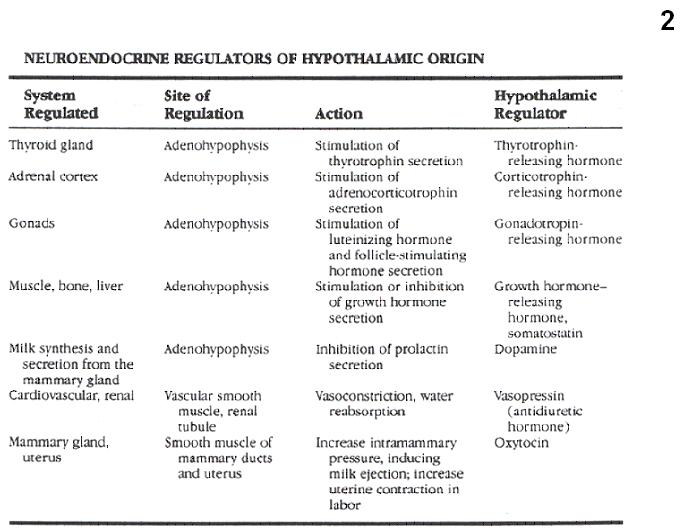{kind=link}
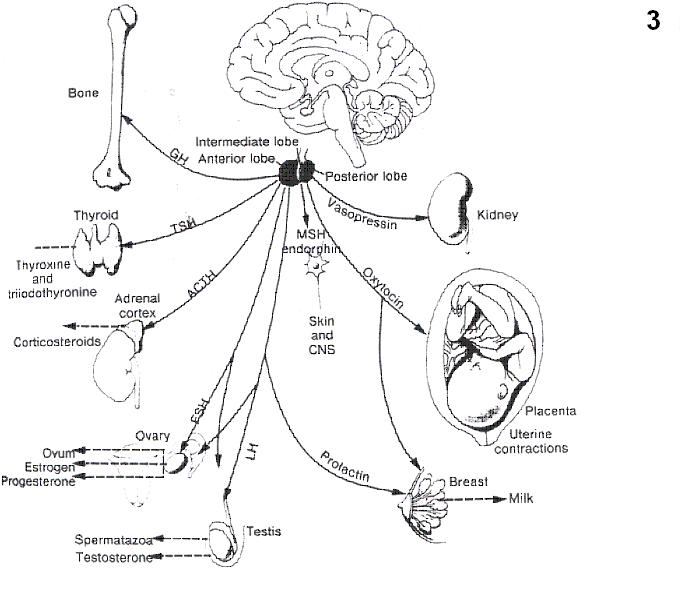{kind=link}
{kind=link}
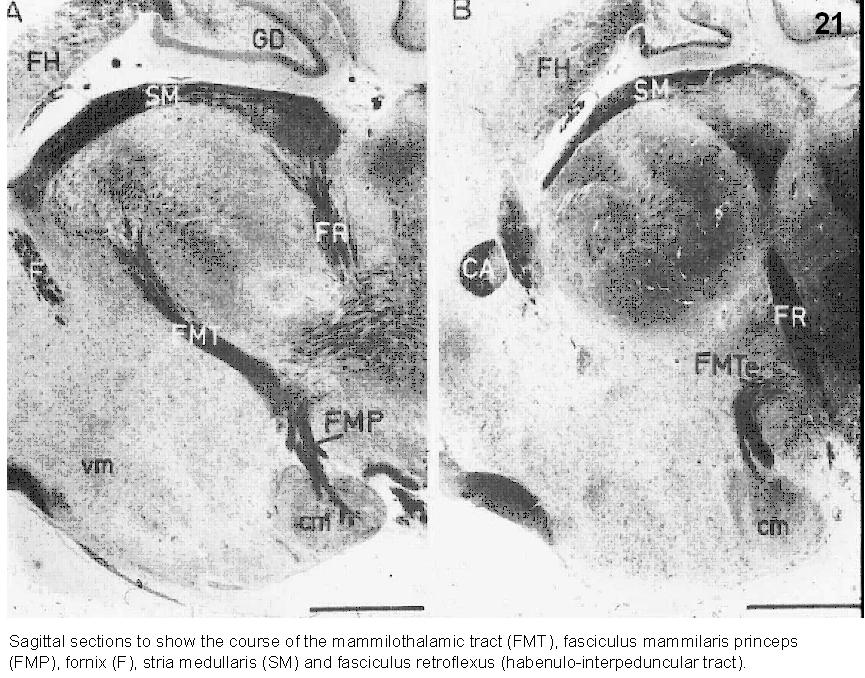{kind=link}
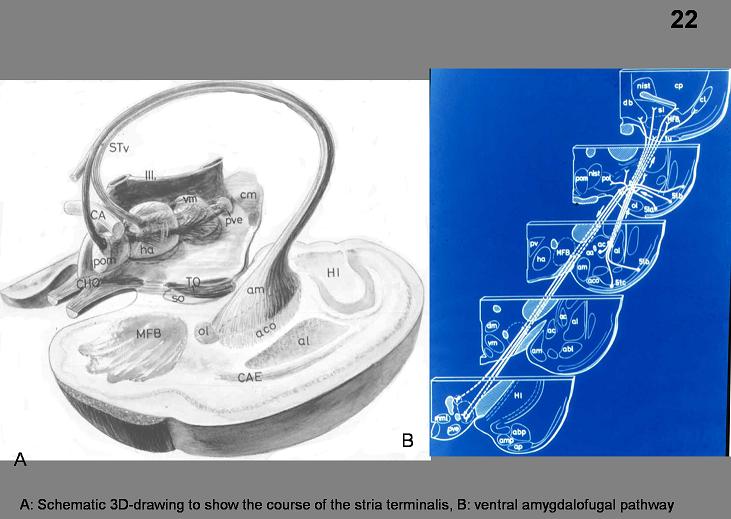{kind=link}
{kind=link}
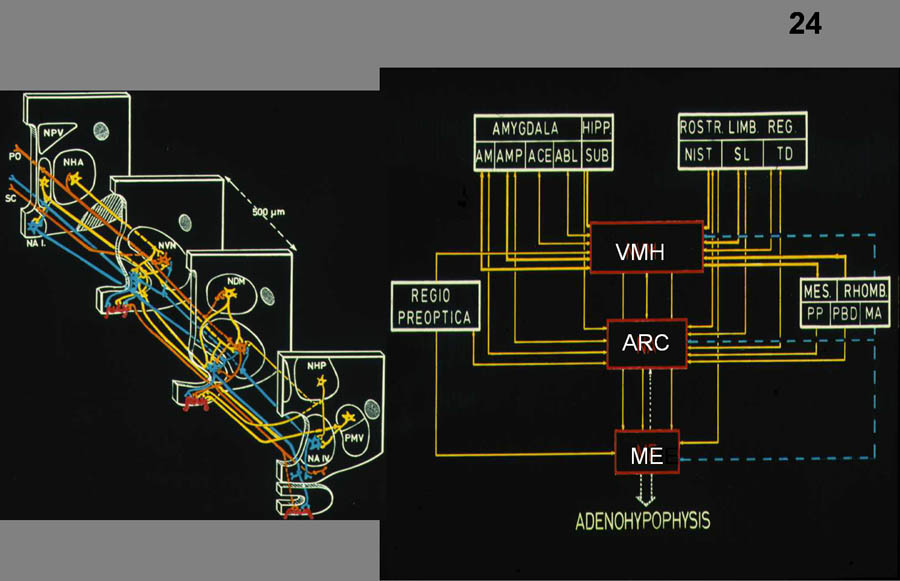{kind=link}
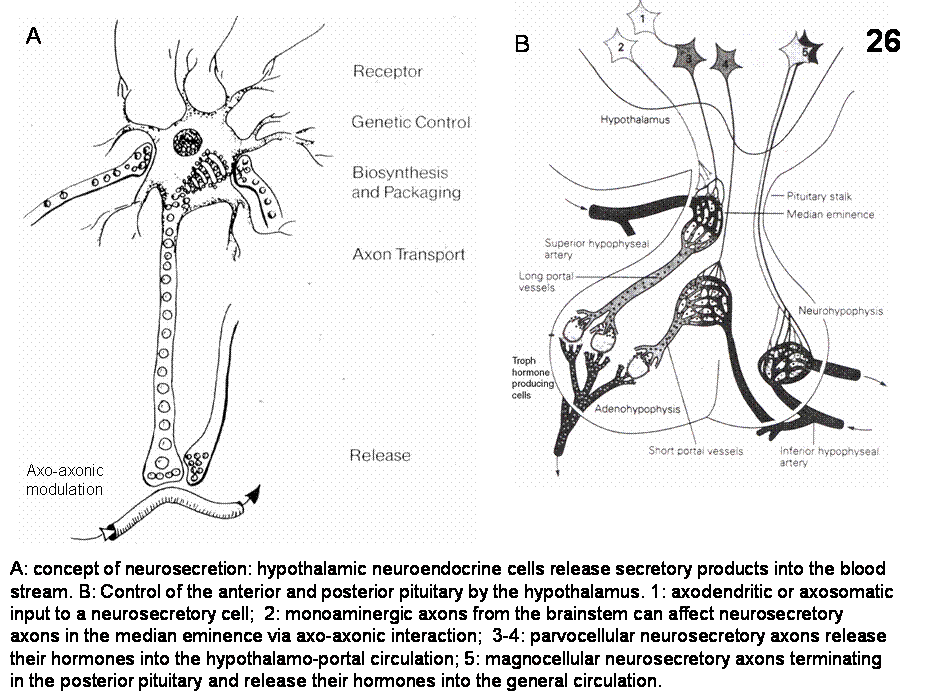{kind=link}
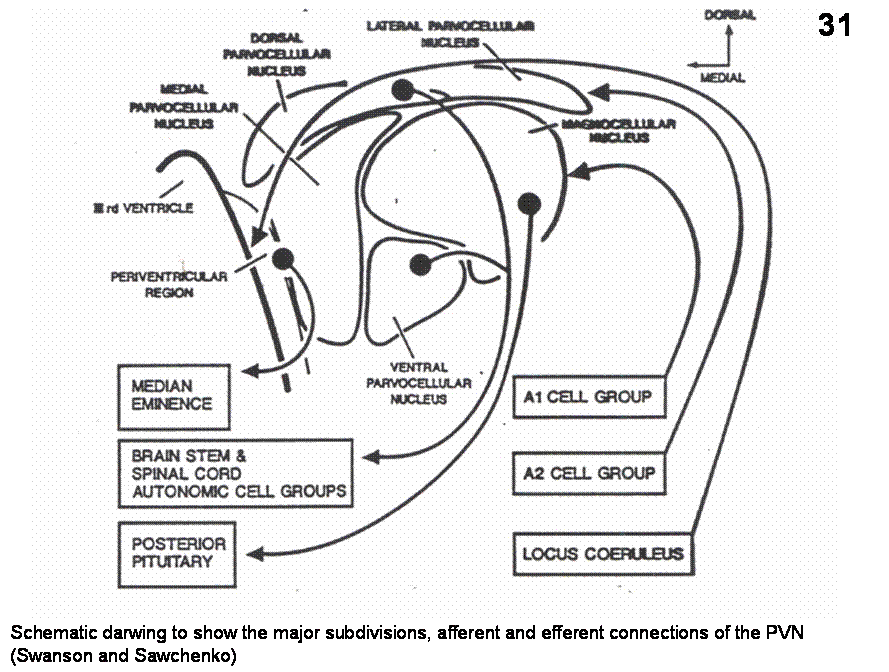{kind=link}
{kind=link}
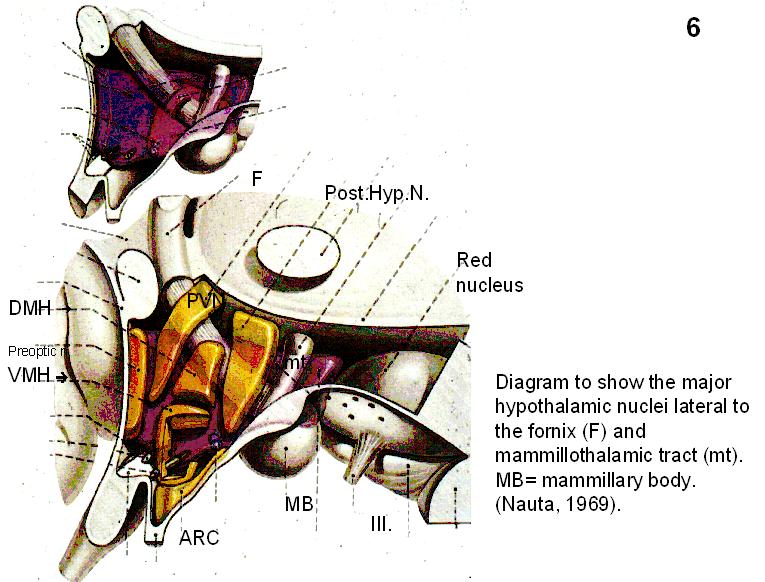{kind=link}
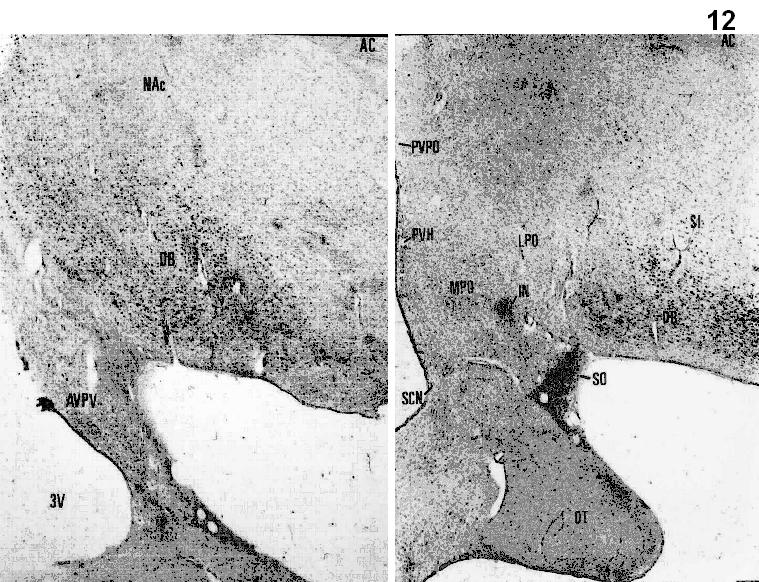{kind=link}
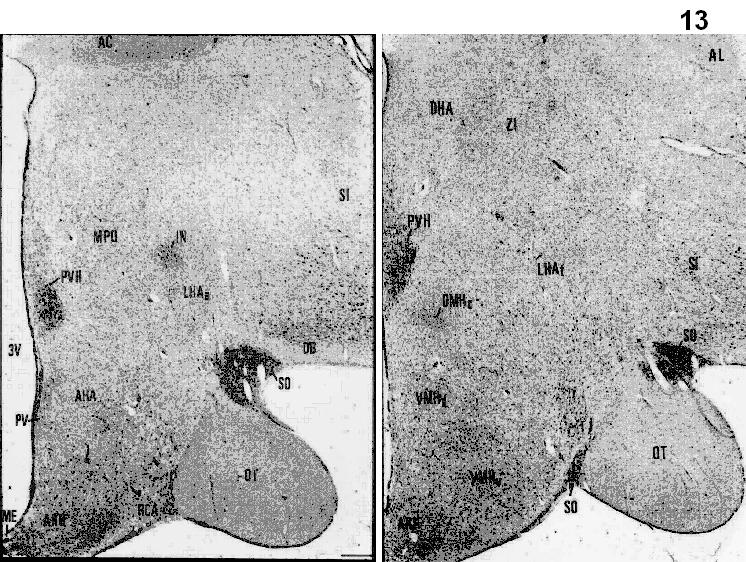{kind=link}
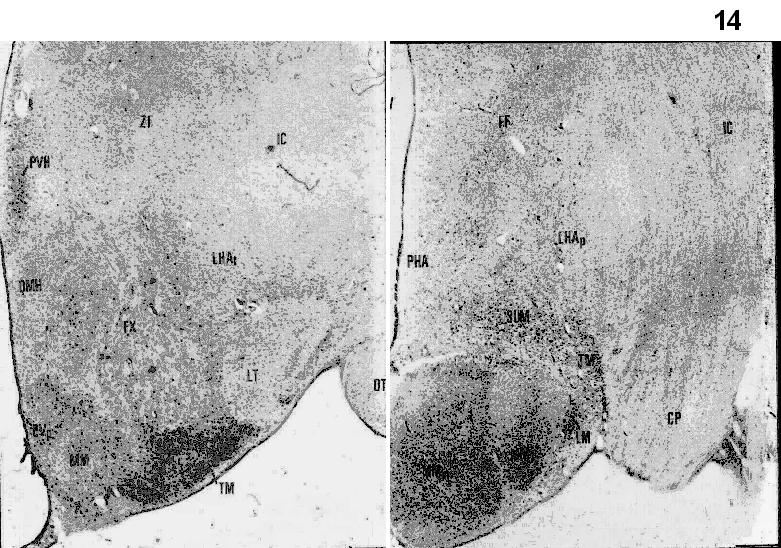{kind=link}
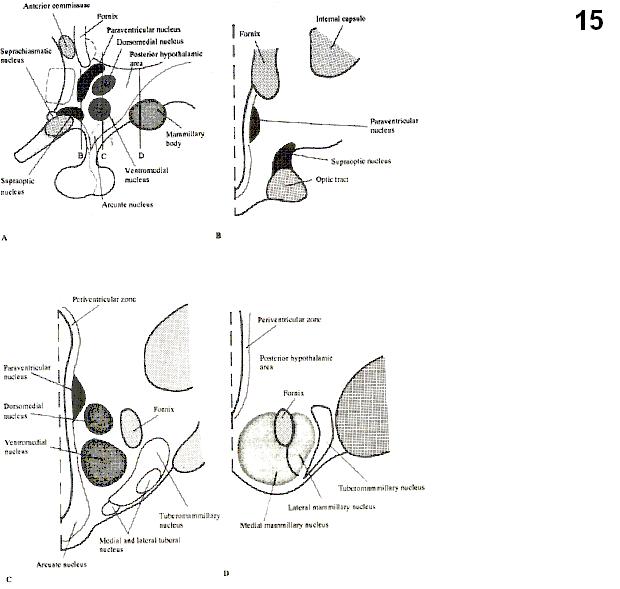{kind=link}
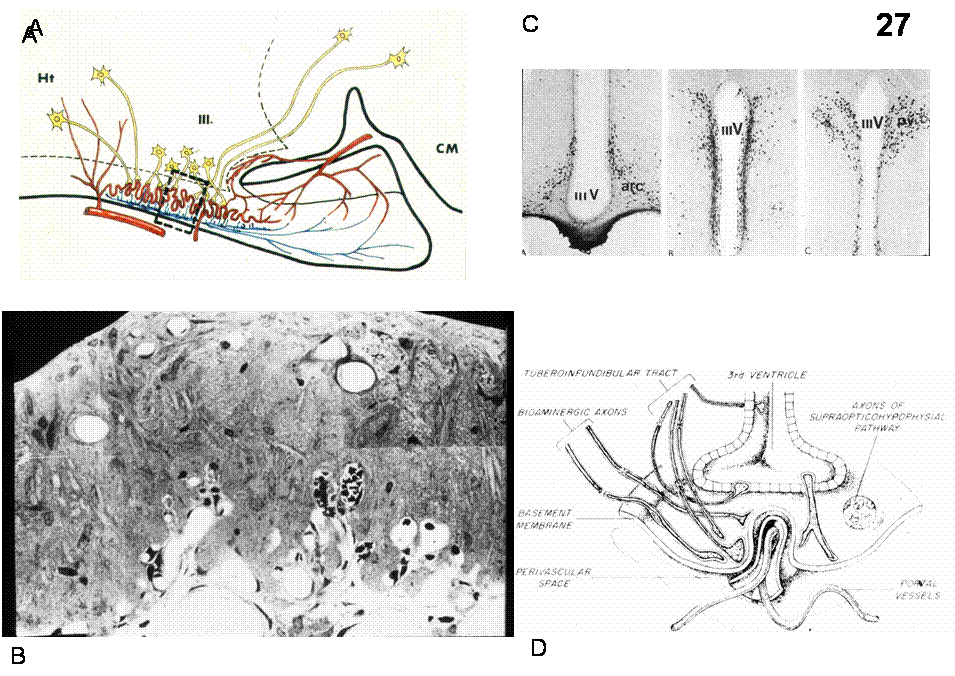{kind=link}
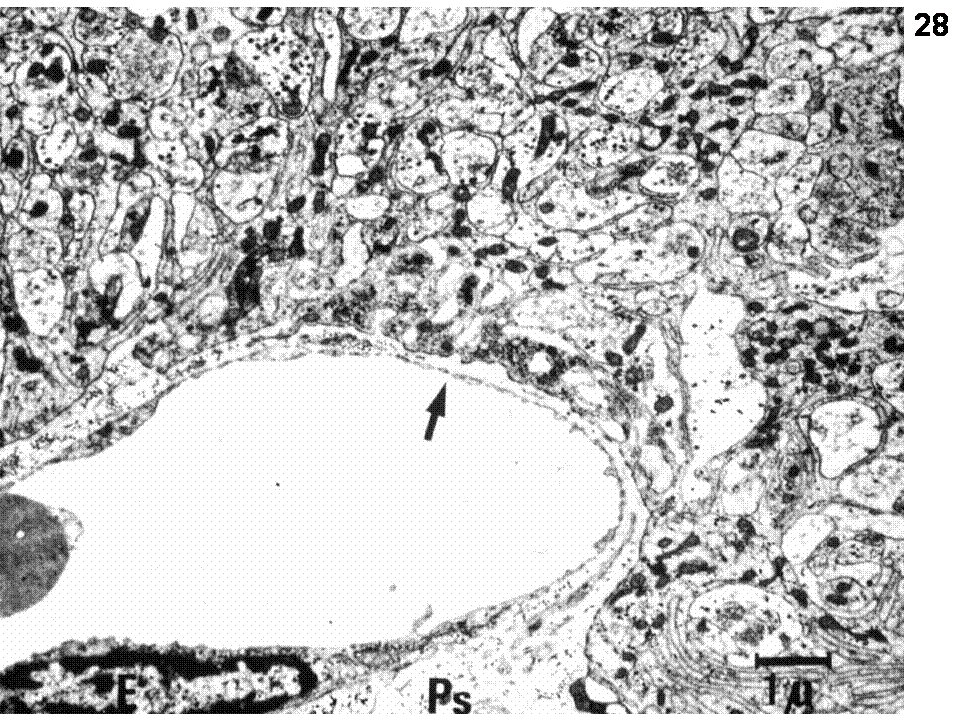{kind=link}
{kind=link}
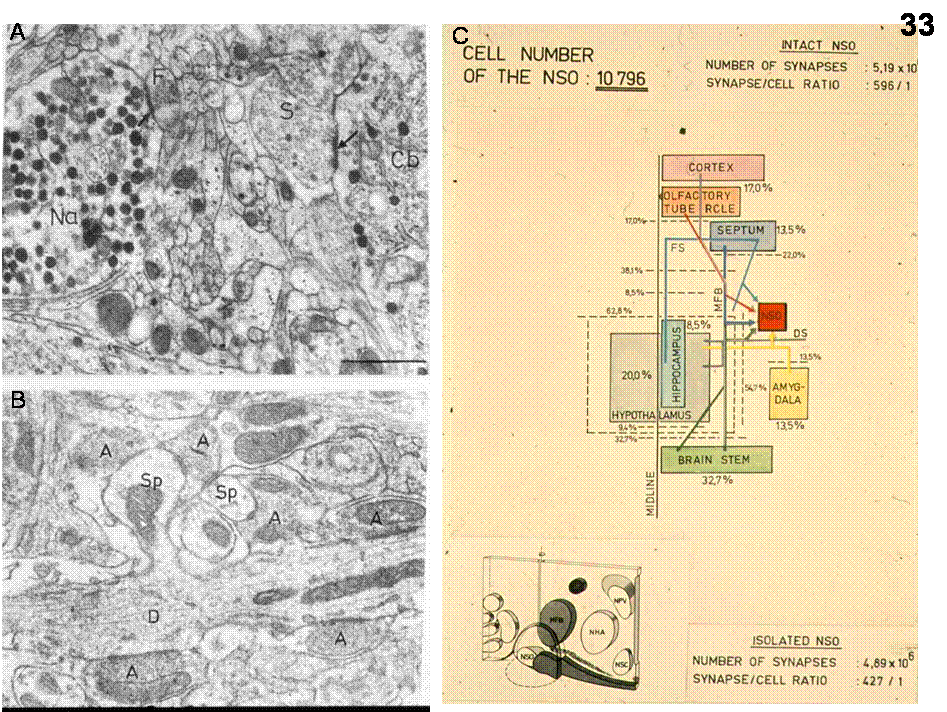{kind=link}
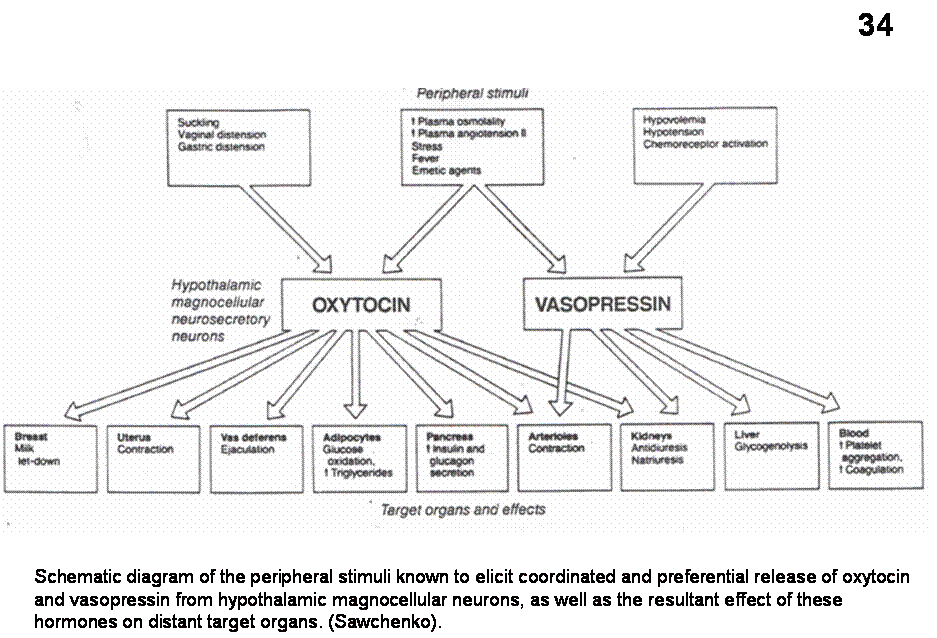{kind=link}
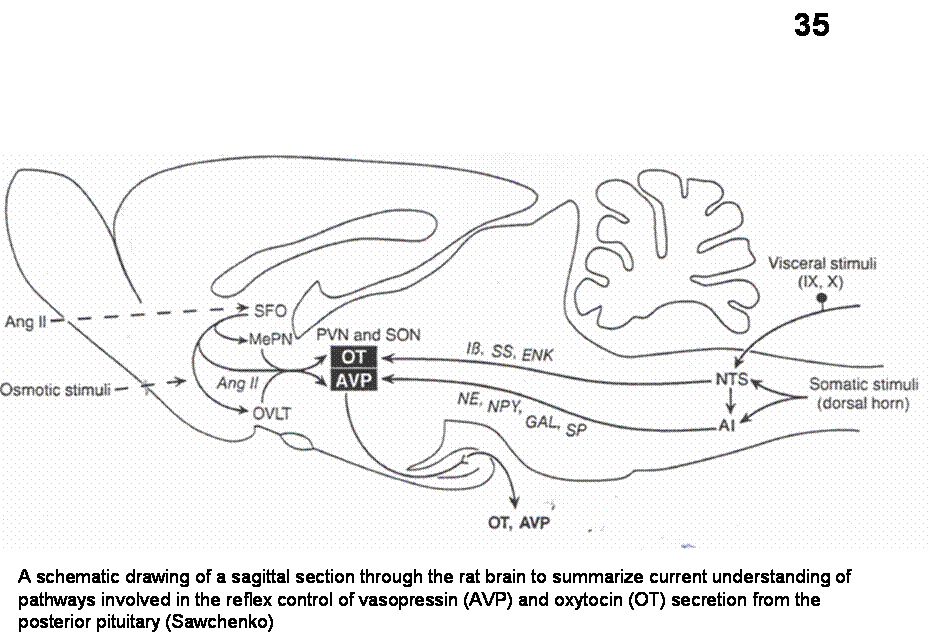{kind=link}
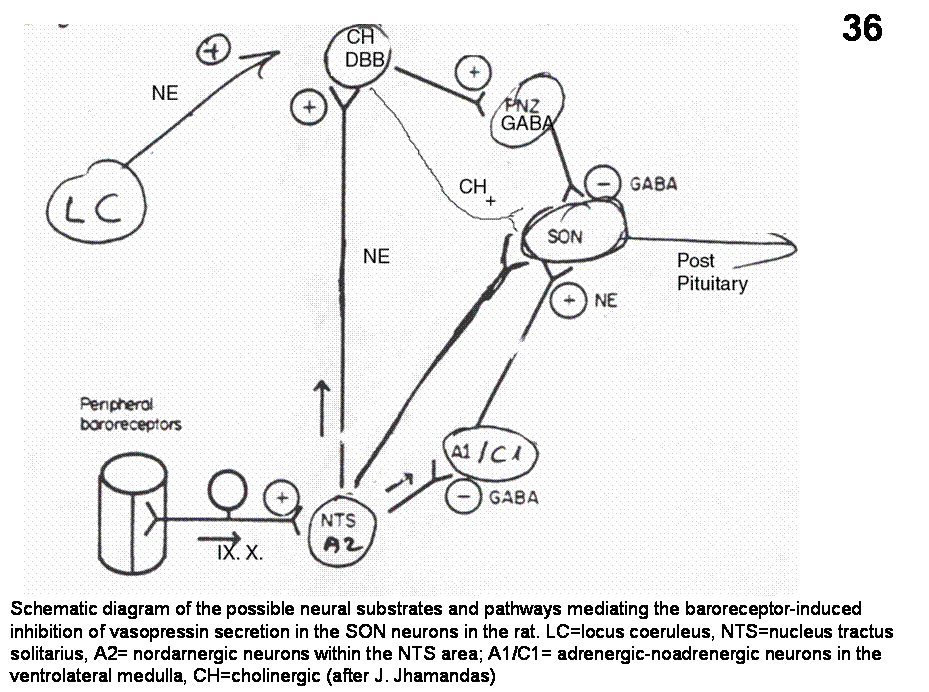{kind=link}
{kind=link}
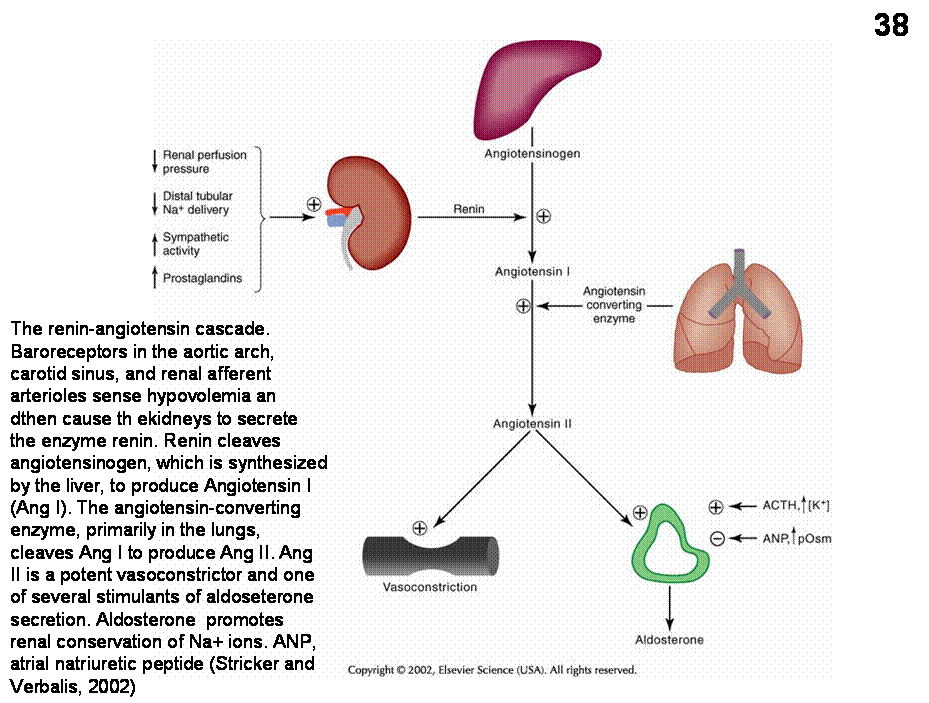{kind=link}
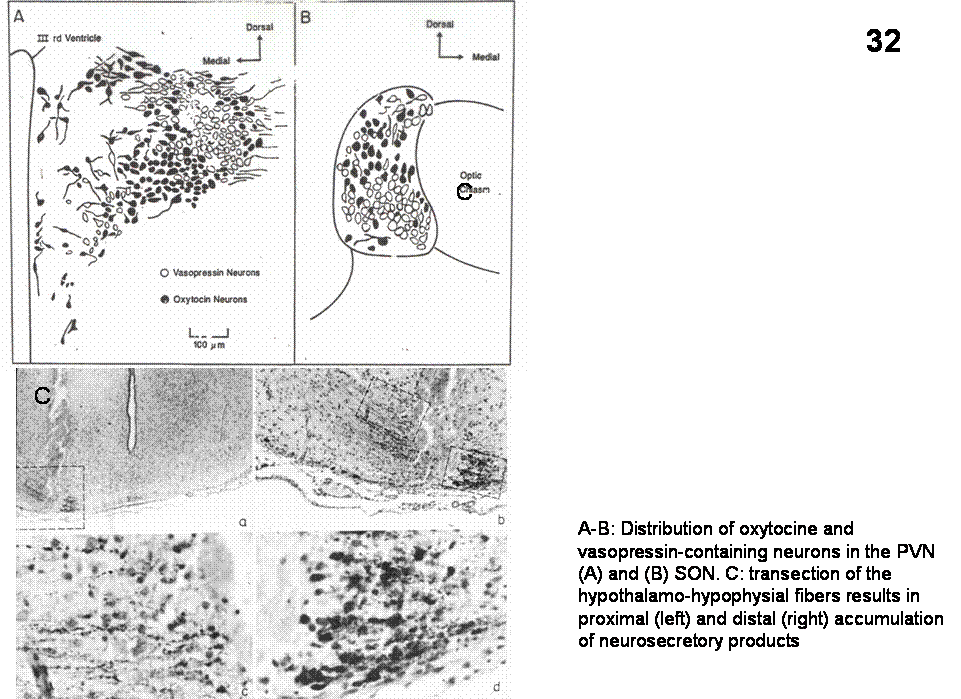{kind=link}
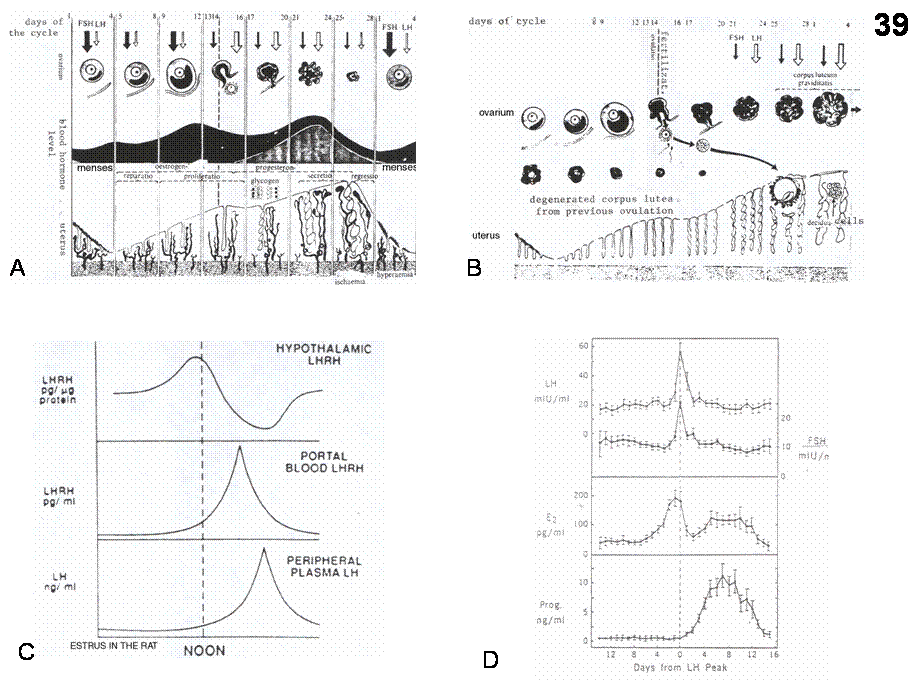{kind=link}
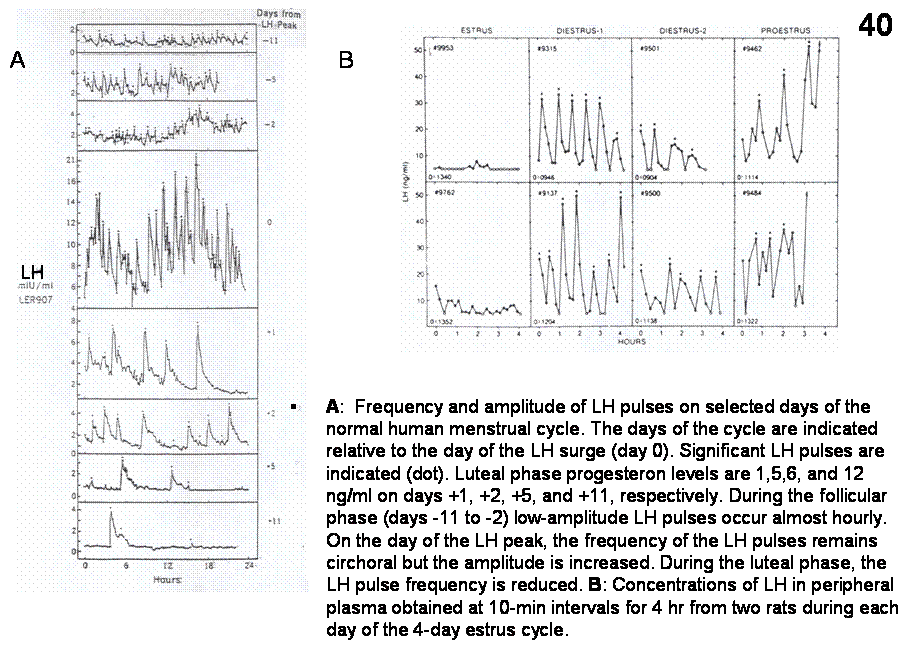{kind=link}
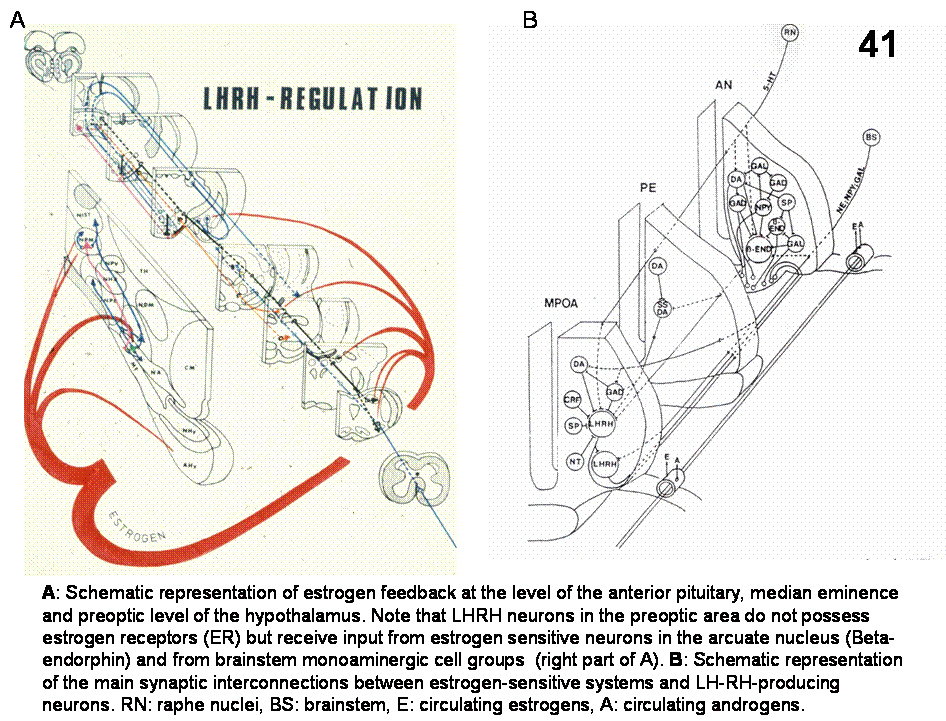{kind=link}
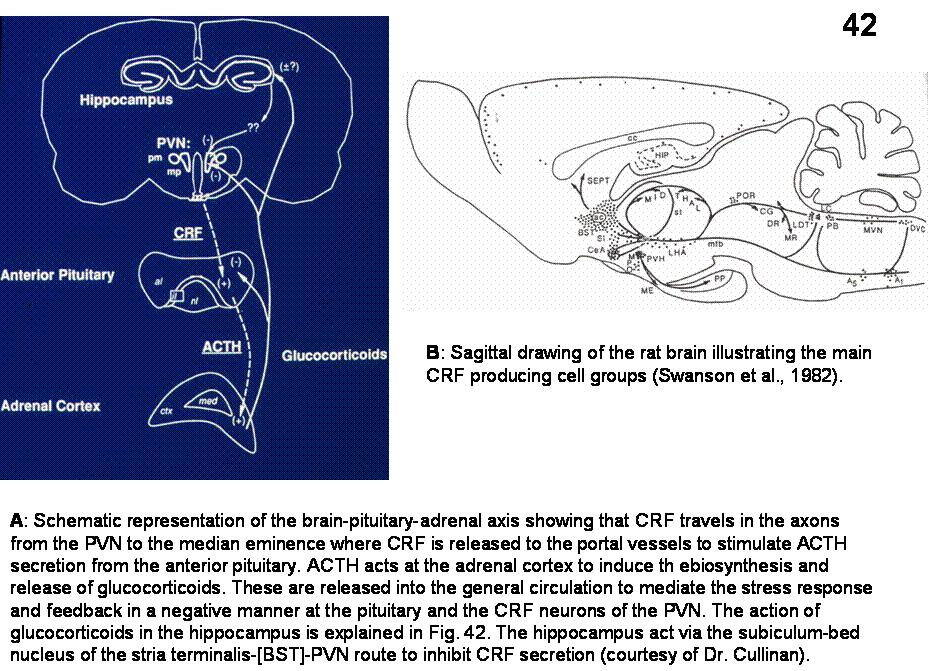{kind=link}
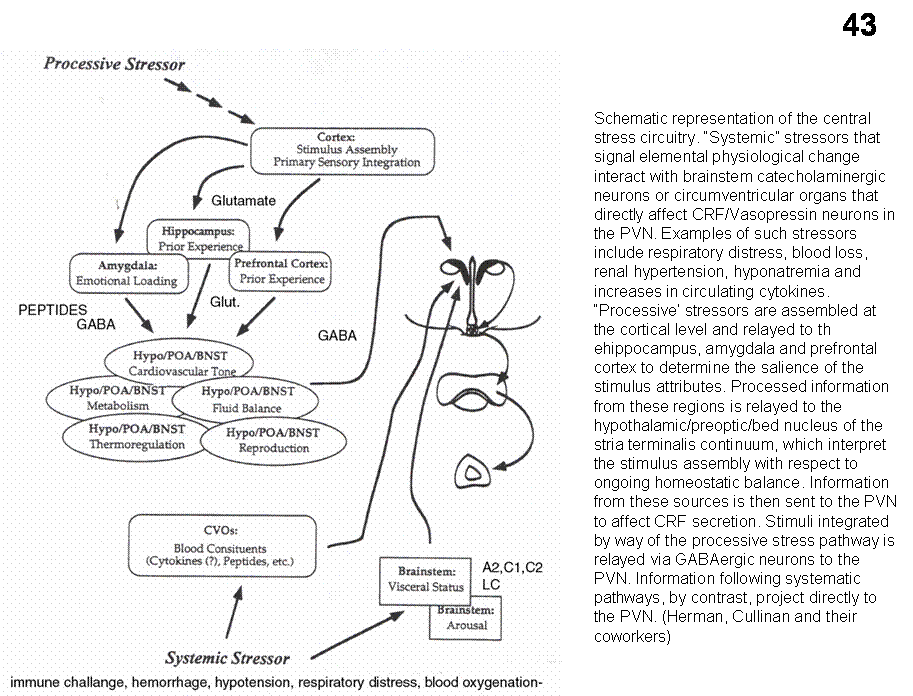{kind=link}
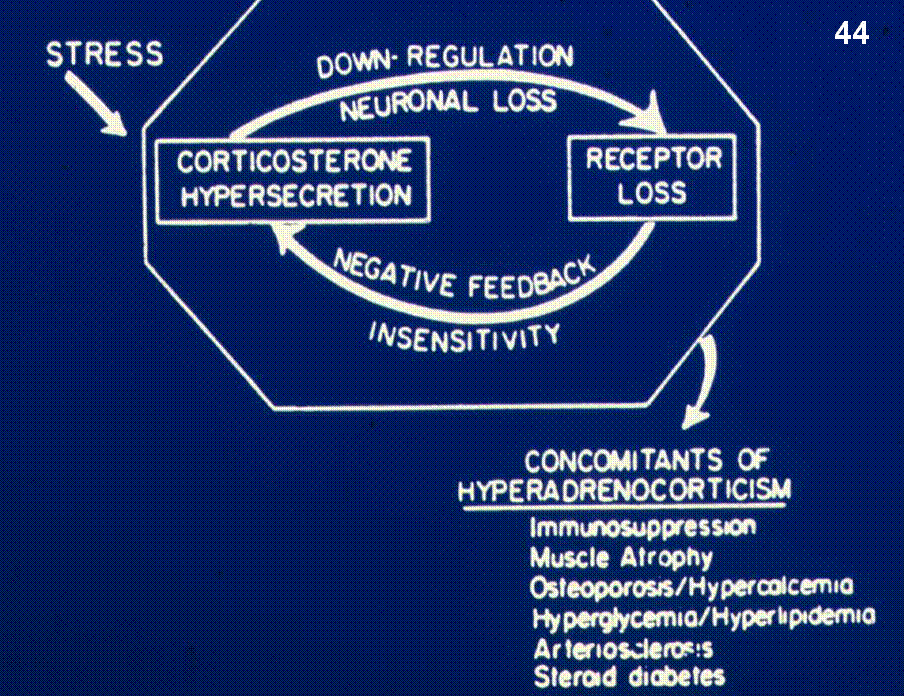{kind=link}
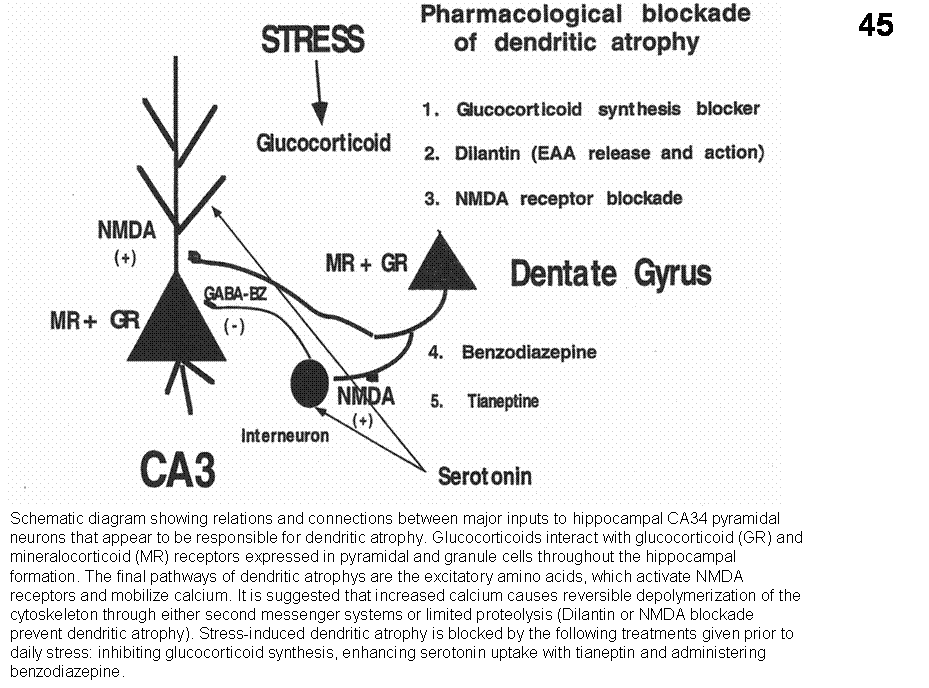{kind=link}
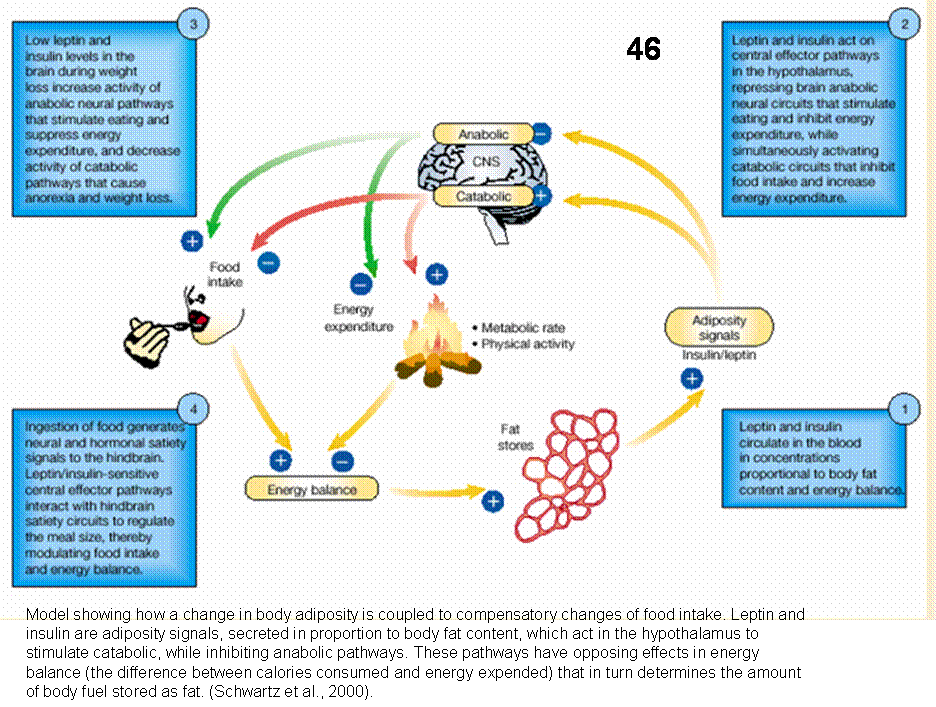{kind=link}
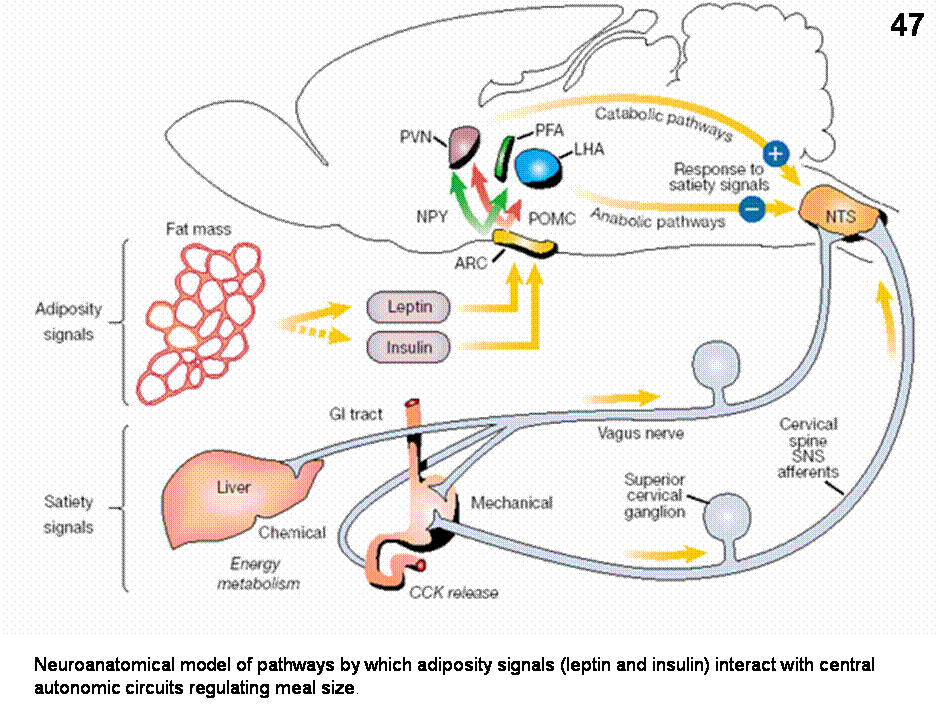{kind=link}
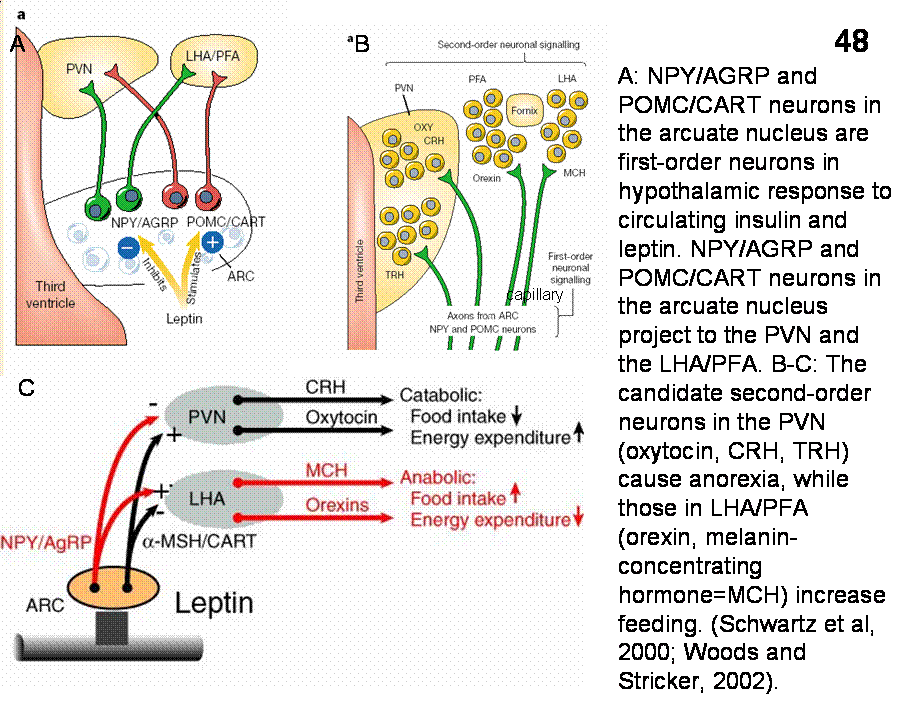{kind=link}
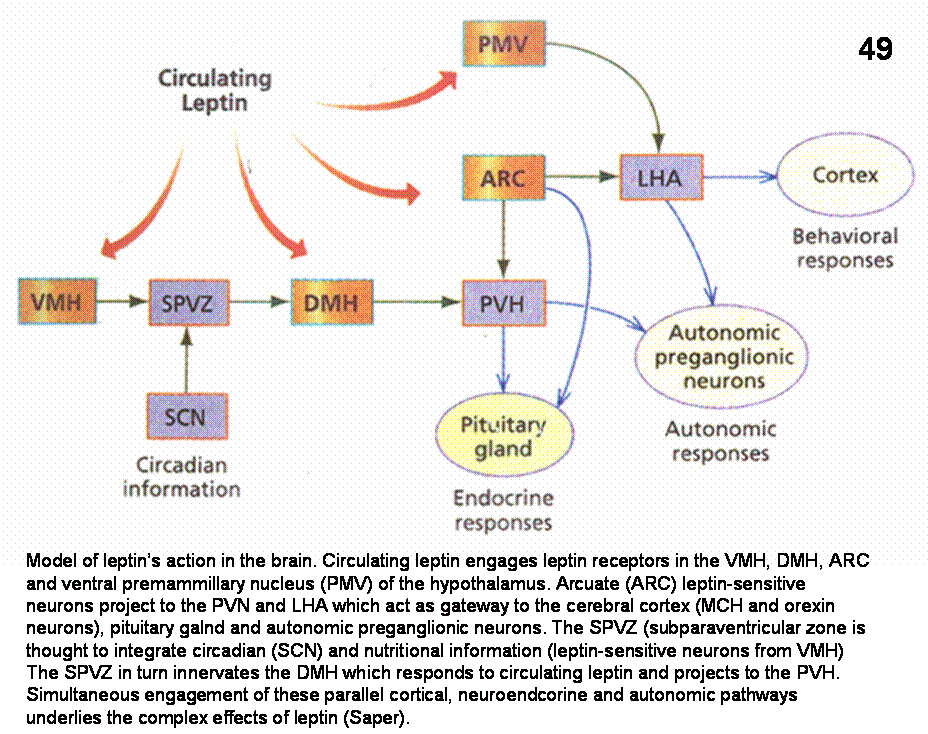{kind=link}
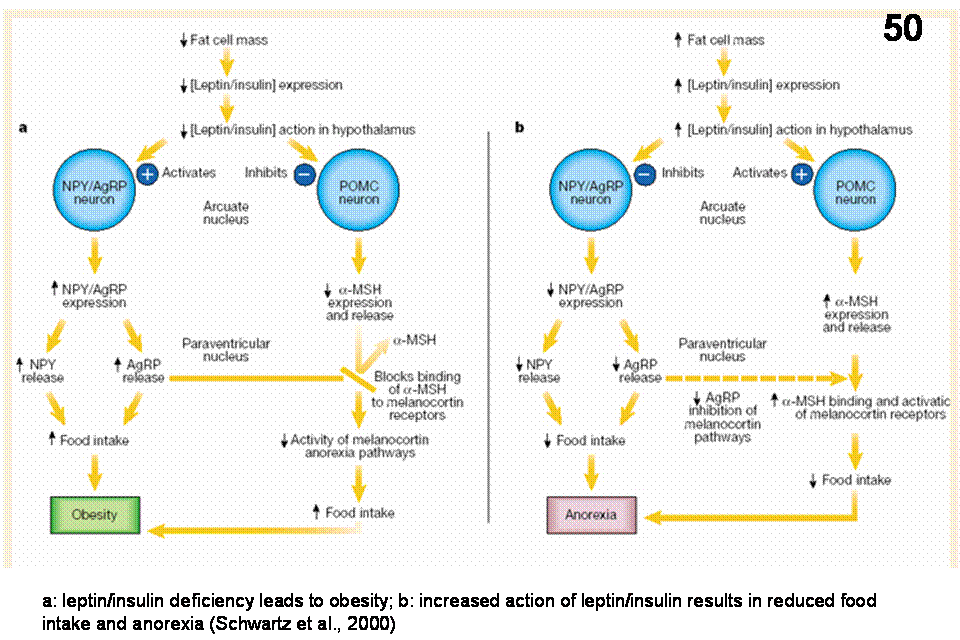{kind=link}
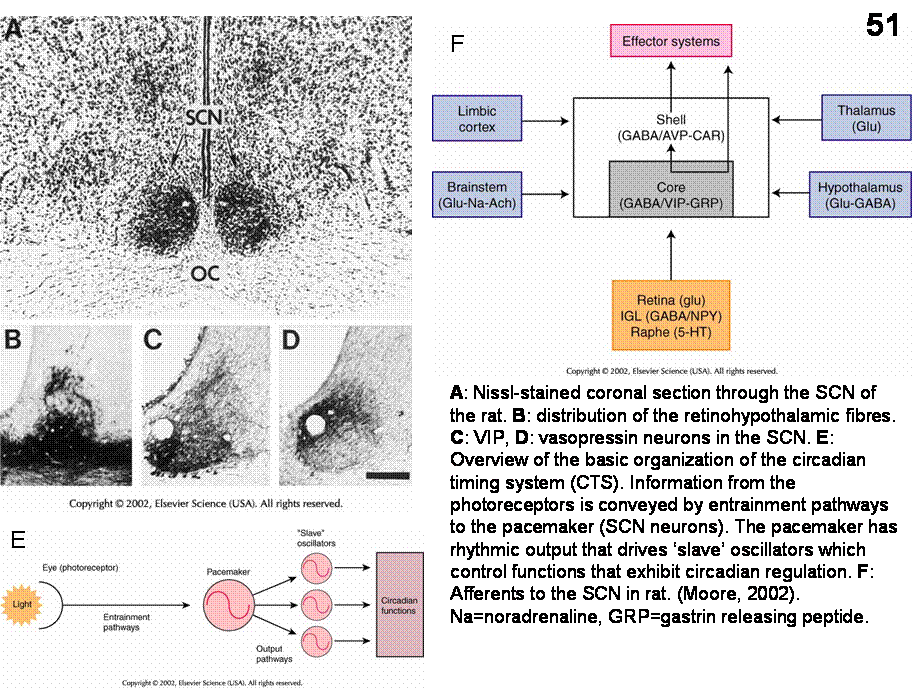{kind=link}
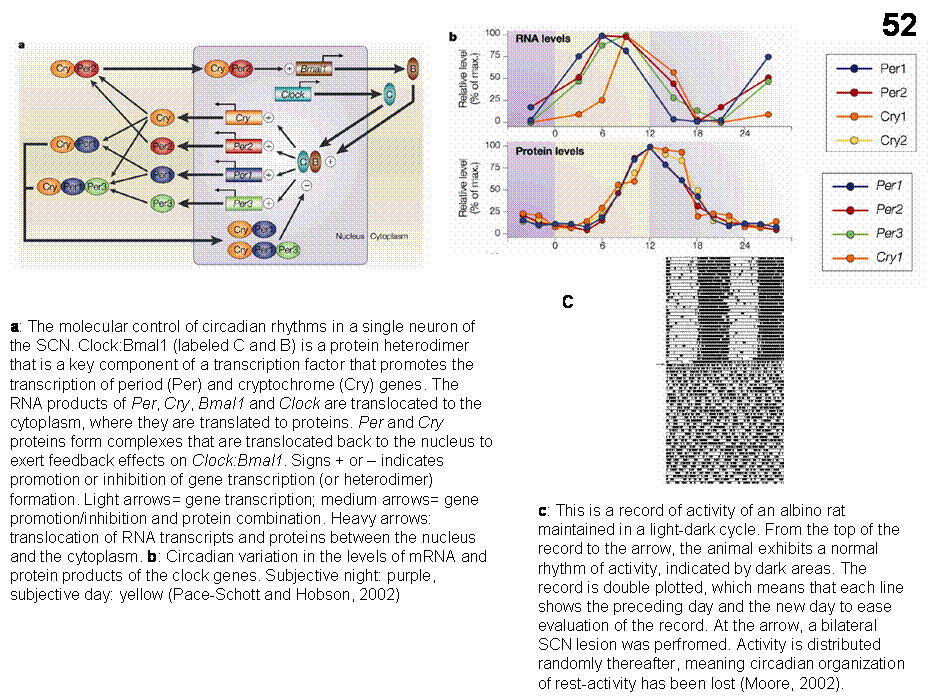{kind=link}
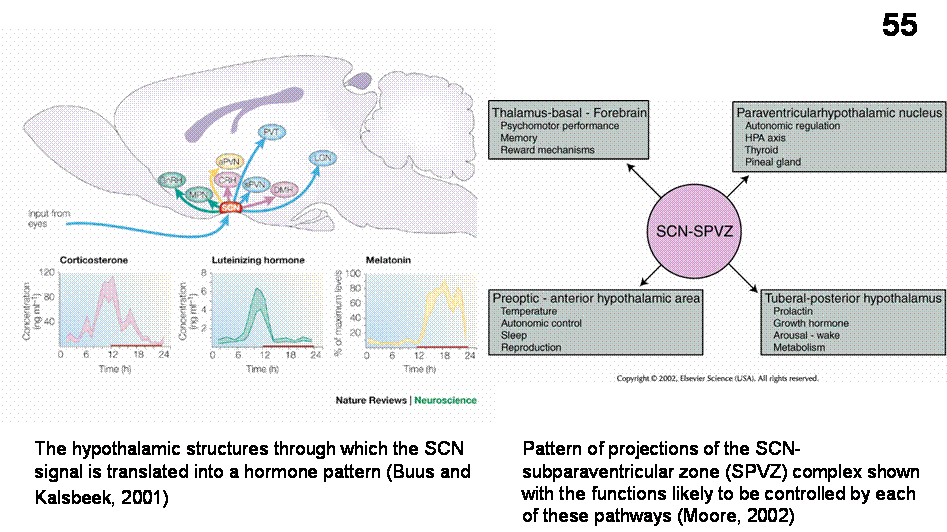{kind=link}
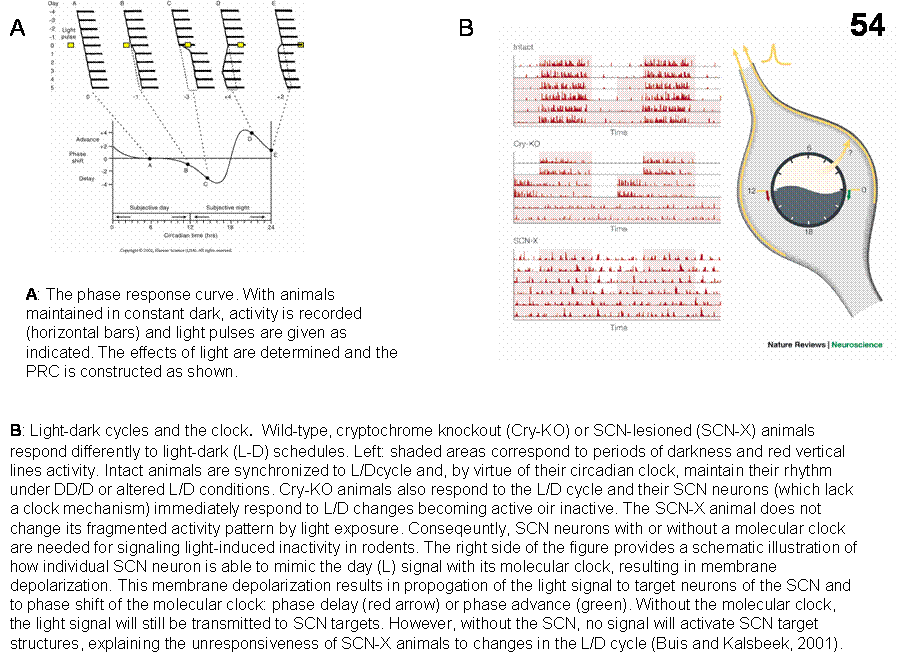{kind=link}
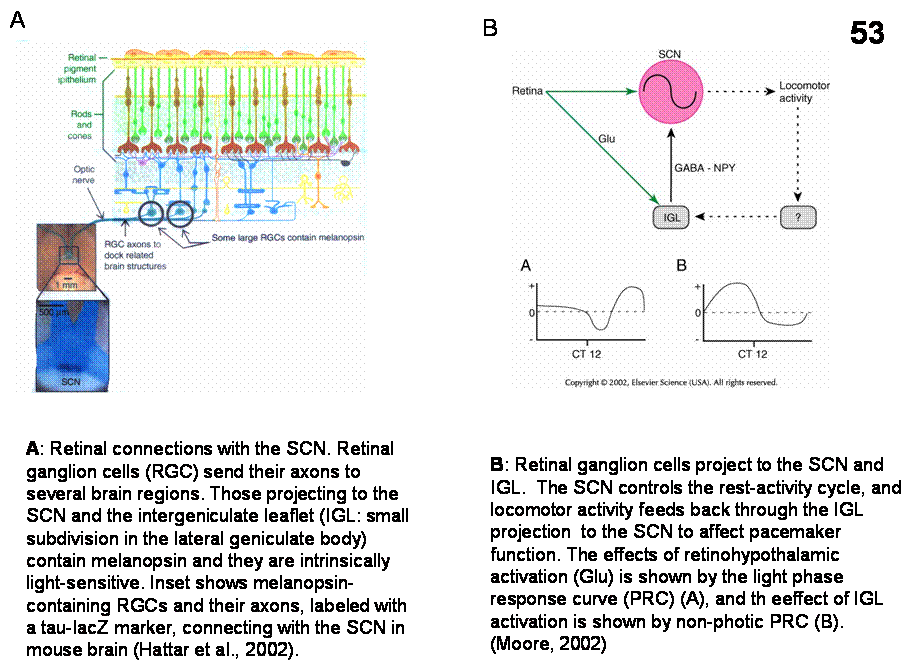{kind=link}
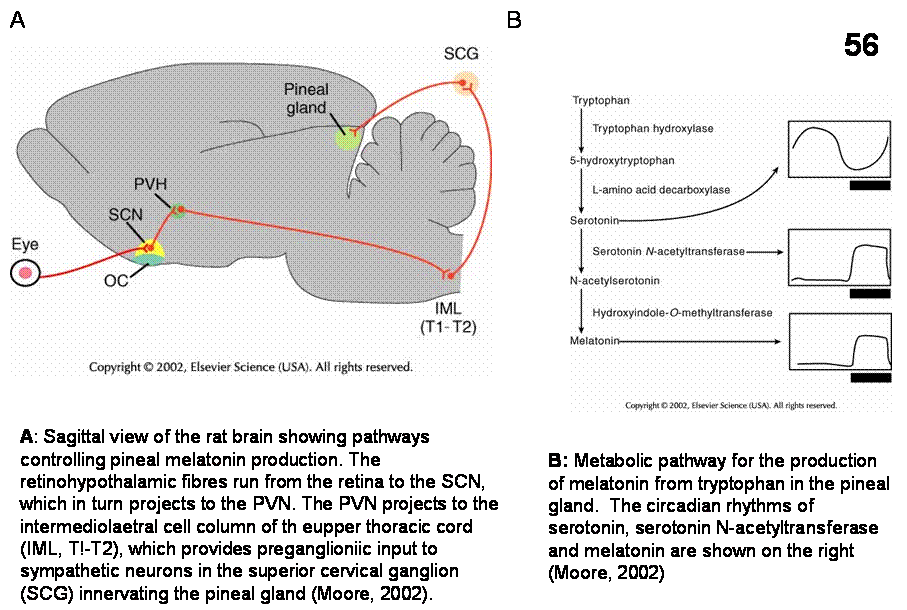{kind=link}
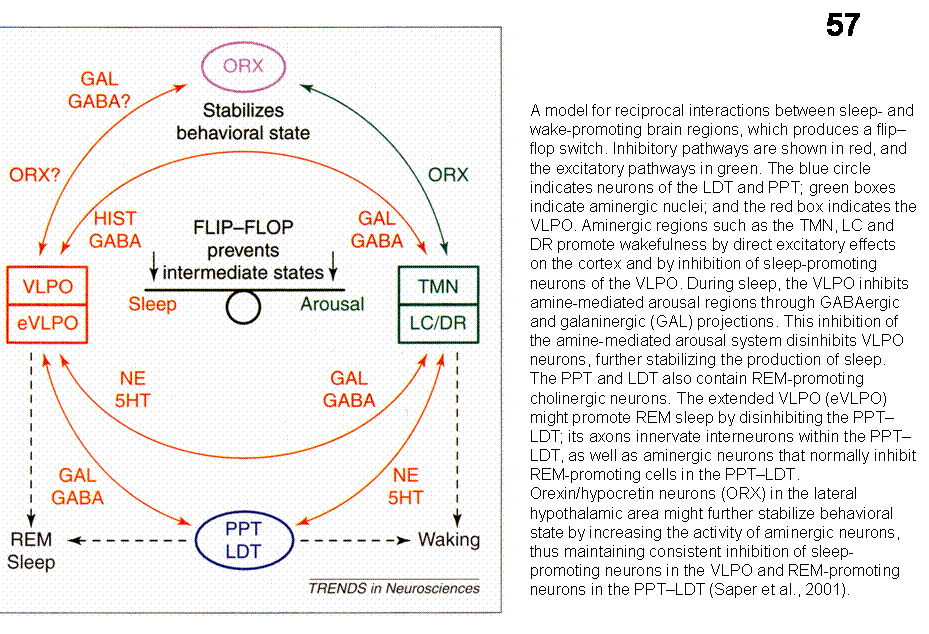{kind=link}
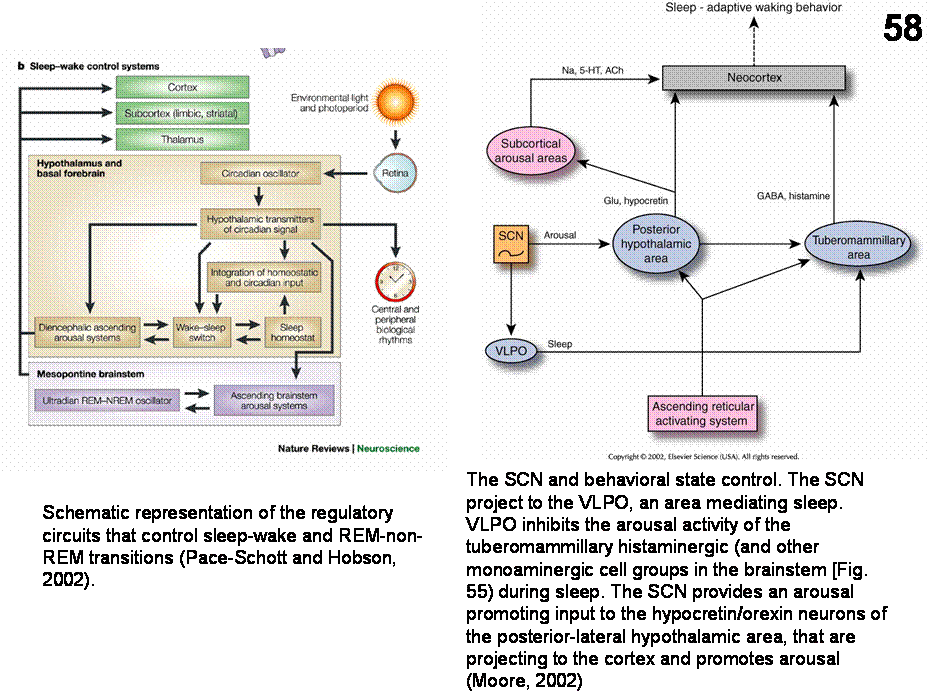{kind=link}